2024-11-08

期刊:Developmental Cell
影響因子:10.7
研究背景
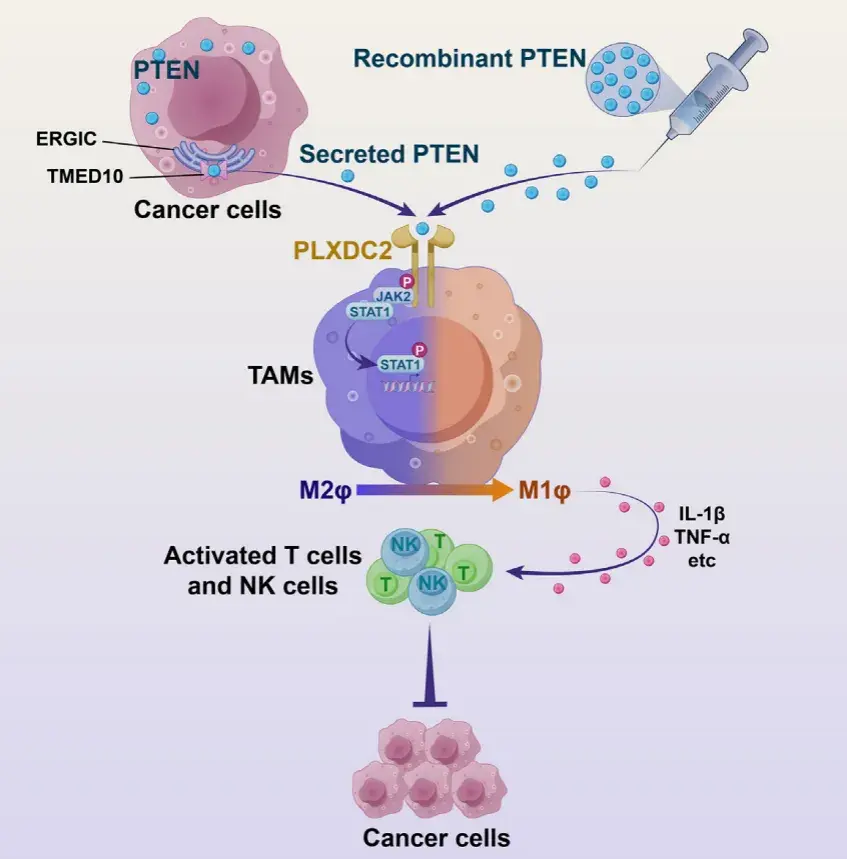
PTEN 是人類癌癥中最常見的腫瘤抑制因子之一,其功能的缺失與多種癌癥的發生、發展以及對免疫治療的耐藥性密切相關。然而,已有研究主要集中在 PTEN 的細胞內功能,對其在腫瘤微環境(TME)中的免疫調節作用了解有限。特別是,PTEN 缺失的腫瘤通常表現出免疫抑制微環境,增加了腫瘤對免疫療法的抵抗力。近年來,研究表明 PTEN 可以通過一種非傳統的蛋白質分泌途徑分泌到細胞外,然而其在腫瘤免疫中的具體機制尚未明確。2024年8月27日,上海交通大學醫學院附屬仁濟醫院/海南醫科大學陳國強院士團隊在Developmental Cell上發表了文章Secreted PTEN binds PLXDC2 on macrophages to drive antitumor immunity and tumor suppression,揭示了PTEN可通過非經典蛋白分泌通路THU以可溶形式分泌到細胞外,并通過重編程腫瘤免疫微環境,實現抗腫瘤免疫的激活與腫瘤抑制。派森諾生物為該研究提供了單細胞轉錄組測序服務。
結 果
01 、PTEN 通過 TMED10 通道蛋白分泌途徑分泌
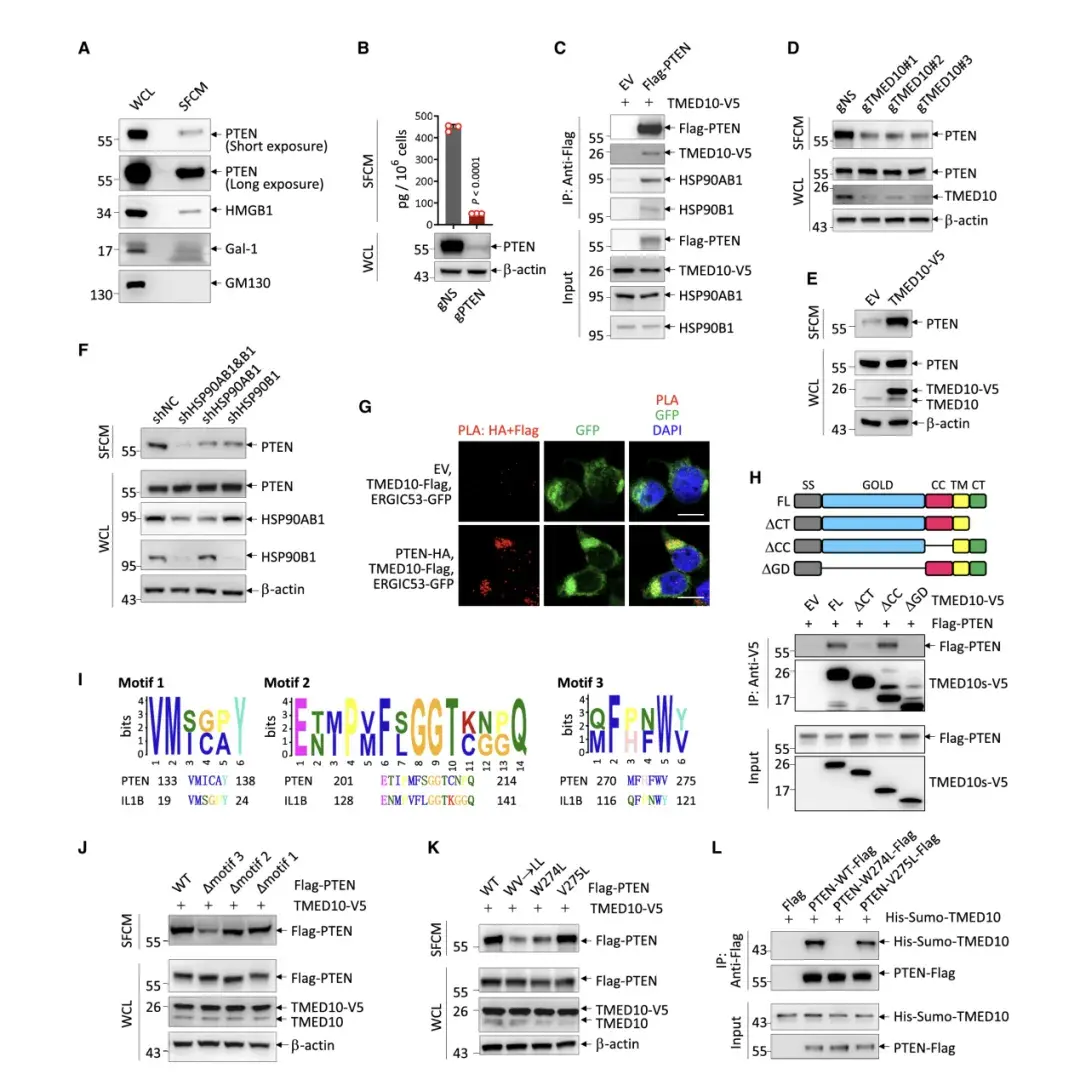
作者在多種細胞系的培養上清液中都發現了PTEN蛋白及其他已知的分泌蛋白,如HMGB1 和Gal-1。而293T 細胞上清液中缺少GM130,這排除了上清液中的PTEN蛋白來自死細胞的可能性。另一方面,從PTEN 敲除的小鼠胚胎成纖維細胞和PTEN缺陷的U251膠質母細胞瘤細胞上清液中未檢測到PTEN,進一步證明 PTEN來自活細胞。ELISA試驗表明,293T細胞在1小時內分泌了450 pg PTEN ,而通過CRISPR-Cas9 系統敲除 PTEN 則消除了該分泌。這些結果表明,細胞內的 PTEN 蛋白可被分泌到細胞外。
考慮到有報道稱 PTEN 是通過外泌體分泌的,作者還探討了外泌體介導 PTEN 分泌的可能性。為此,作者收集了與293T細胞孵育1小時和24小時的培養基,通過超濾分離和TEM分析,發現外泌體中PTEN的含量有限,證明外泌體不是 PTEN 分泌的主要途徑。
然后作者進一步排除了 PTEN 通過常規蛋白質分泌途徑分泌的可能性,因為布雷菲德菌素 A 可阻斷分泌蛋白從內質網到高爾基體的運輸,但對293T 細胞分泌內源性 PTEN 沒有影響。這與使用 SignalP 預測的 PTEN 蛋白缺乏典型分泌信號的事實一致。為了找到 PTEN 分泌途徑的線索,作者轉向 PTEN 相互作用蛋白,發現PTEN與HSP90B1、TMED10和HSP90AB1相互作用,這些是THU通路的組成部分。敲除 TMED10發現可顯著降低分泌的 PTEN 量,而過表達 TMED10 可進一步增加分泌的 PTEN 量。此外,HSP90AB1 或 HSP90B1 的耗竭會抑制 PTEN 的分泌,而它們的共同耗竭則顯示出最強的效果。這些結果表明 THU 可能是 PTEN 分泌的主要途徑。
為了鞏固 PTEN 通過 THU 分泌的觀點,作者發現PTEN與TMED10共定位在ERGIC上,并與TMED10中的類似區域結合,如同IL-1b。通過MEME-Suite網站分析,作者發現PTEN和IL-1b有共同基序,其中基序3對PTEN分泌至關重要。W274是指導THU分泌PTEN的關鍵氨基酸殘基,W274突變幾乎消除了PTEN與TMED10的相互作用。PTEN的W274在物種中高度保守。綜上,TMED10通過識別PTEN的W274來指導其通過THU通路。
02、細胞外可溶性 PTEN 抑制腫瘤生長和免疫反應
2.1 細胞外可溶性 PTEN 抑制腫瘤生長
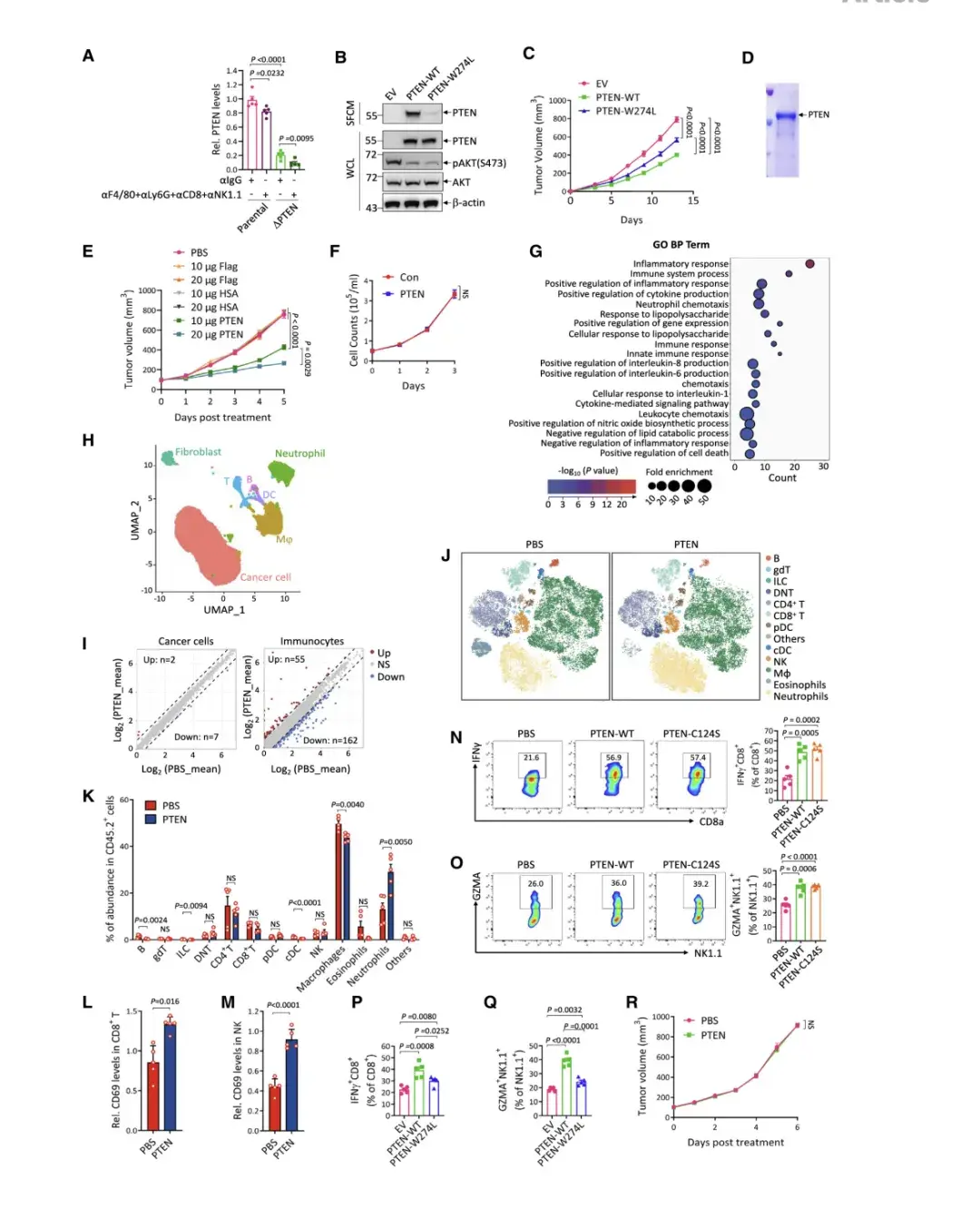
作者用ELISA試驗檢測B16-F10小鼠黑色素瘤細胞腫瘤間質液(TIF)中的PTEN蛋白,發現PTEN以22 ng / mL的濃度存在于TIF中,這驗證了TME中細胞外PTEN的存在。而后為了確定TIF中PTEN的來源,比較親本B16-F10細胞和PTEN敲除的B16-F10細胞系產生的TIF,發現后者中PTEN減少了約80%,說明腫瘤細胞是TME中分泌PTEN的主要來源。
為了評估分泌型 PTEN 在 TME 中的作用,將 PTEN 敲除的 MC38 細胞 (MC38-DPTEN)進行野生型 PTEN (PTEN-WT) 或含有 W274L 突變的 PTEN 突變體 (PTEN-W274L)再表達,發現PTEN-W274L 的分泌效率低于 PTEN-WT,但它們對 AKT 磷酸化 (Ser473) 的抑制強度相似 ,這表明 W274L 突變選擇性地影響 PTEN 的分泌,但不影響其磷酸酶活性。然后,將這些細胞系在 C57BL/6J 小鼠中進行皮下生長,結果顯示與 PTEN-WT 相比,PTEN W274L 的腫瘤抑制作用受損 ,這表明分泌型 PTEN 可能在腫瘤抑制中發揮作用
為了進一步評估分泌型 PTEN 在腫瘤抑制中的作用,作者嘗試直接用 PTEN 蛋白治療腫瘤。
與所有對照組相比,PTEN 蛋白治療表現出顯著的劑量依賴性腫瘤抑制作用。細胞外 PTEN 的腫瘤抑制作用也可以在 MC38-DPTEN 細胞產生的腫瘤中重現。值得注意的是,純化的 PTEN 蛋白攜帶一種使其磷酸酶活性失活的突變(PTEN-C124S),其對腫瘤生長的抑制程度與 PTEN-WT 相似。此外,腹膜內注射純化的 PTEN 也顯著抑制了B16-F10-DPTEN 腫瘤的生長。總之,細胞外可溶性PTEN以磷酸酶活性非依賴的方式發揮腫瘤抑制作用。
2.2 細胞外 PTEN 引發抗腫瘤免疫反應
與體內抑瘤作用相反,細胞外 PTEN 治療不影響 B16-F10DPTEN 細胞的體外生長,這暗示它可能作用于TME。事實上,當通過 RNA 測序分析 PTEN 治療的 B16-F10-DPTEN 腫瘤時,對 104 個失調基因的 GO 分析顯示,它們主要富集在參與免疫反應的途徑中。特別是,干擾素 a 和 IFNg 反應的強烈激活表明 PTEN 治療會在腫瘤中引發抗腫瘤免疫反應。此外,作者對 PTEN 治療和未治療的 B16-F10DPTEN 腫瘤進行單細胞 RNA 轉錄組測序 (scRNA-seq)。結果顯示,與轉錄組幾乎未受干擾的癌細胞相比,免疫細胞的轉錄組發生了顯著變化,表明 PTEN 治療優先改變了 TIME。
接著,作者使用質譜流式細胞術對接受 PTEN 治療和未接受 PTEN 治療的 B16-F10-DPTEN 腫瘤中的免疫細胞進行了分析。中性粒細胞總數變化最為顯著,巨噬細胞總數略有改變,而 CD4+ /CD8+ T 細胞和 NK 細胞的總數未受到影響。然而,在接受 PTEN 治療的腫瘤中,表達活化標志物 CD69 的 CD8+ T 細胞和 NK 細胞的比例顯著增加。通過對 B16-F10DPTEN 腫瘤以及 MC38-DPTEN 腫瘤中的 IFNg 陽性 CD8+ T 細胞和顆粒酶 A (GZMA) 陽性 NK 細胞進行流式細胞術分析,進一步證實了 PTEN-WT 或 PTEN-C124S 治療增強了 CD8+ T 細胞和NK細胞的活化。與此同時,MC38-DPTEN 腫瘤中 PTEN 分泌的喪失導致了免疫抑制表型。結果表明,細胞外 PTEN 可增強抗腫瘤免疫力。此外,當用 PTEN 治療在免疫缺陷 NCG 小鼠中生長的 B16-F10-DPTEN 腫瘤時,未觀察到對腫瘤生長的明顯影響,這表明細胞外 PTEN 以免疫反應依賴的方式抑制腫瘤生長
由于之前已證實 PTEN 無法在沒有遞送劑的幫助下進入細胞,作者推測它是否與 TME 中的免疫細胞表面結合。作者從 PTEN 治療的 B16F10-DPTEN 腫瘤中分離出上述 PTEN 調節的免疫細胞(巨噬細胞、中性粒細胞、T 細胞和 NK 細胞),并測試了 PTEN 在其表面的存在。在四種類型的免疫細胞中,只有巨噬細胞被 PTEN 標記。通過將 PTEN 蛋白與從 C57BL/6J 小鼠脾臟中分離的免疫細胞進行體外孵育,可以確認 PTEN 在巨噬細胞結合中的偏好。此外,PTEN 以劑量依賴性方式與從骨髓來源的巨噬細胞(BMDM)和腹腔中分離的巨噬細胞結合。將 FLAG 標記的 PTEN 與未通透的 BMDM 一起孵育后,通過對 PTEN 進行免疫熒光染色,也可以觀察到 PTEN 在 BMDM 表面上的存在,而未觀察到細胞內信號。所有這些結果都表明 PTEN 與巨噬細胞表面結合。
03、細胞外 PTEN 使巨噬細胞極化為 M1 樣表型
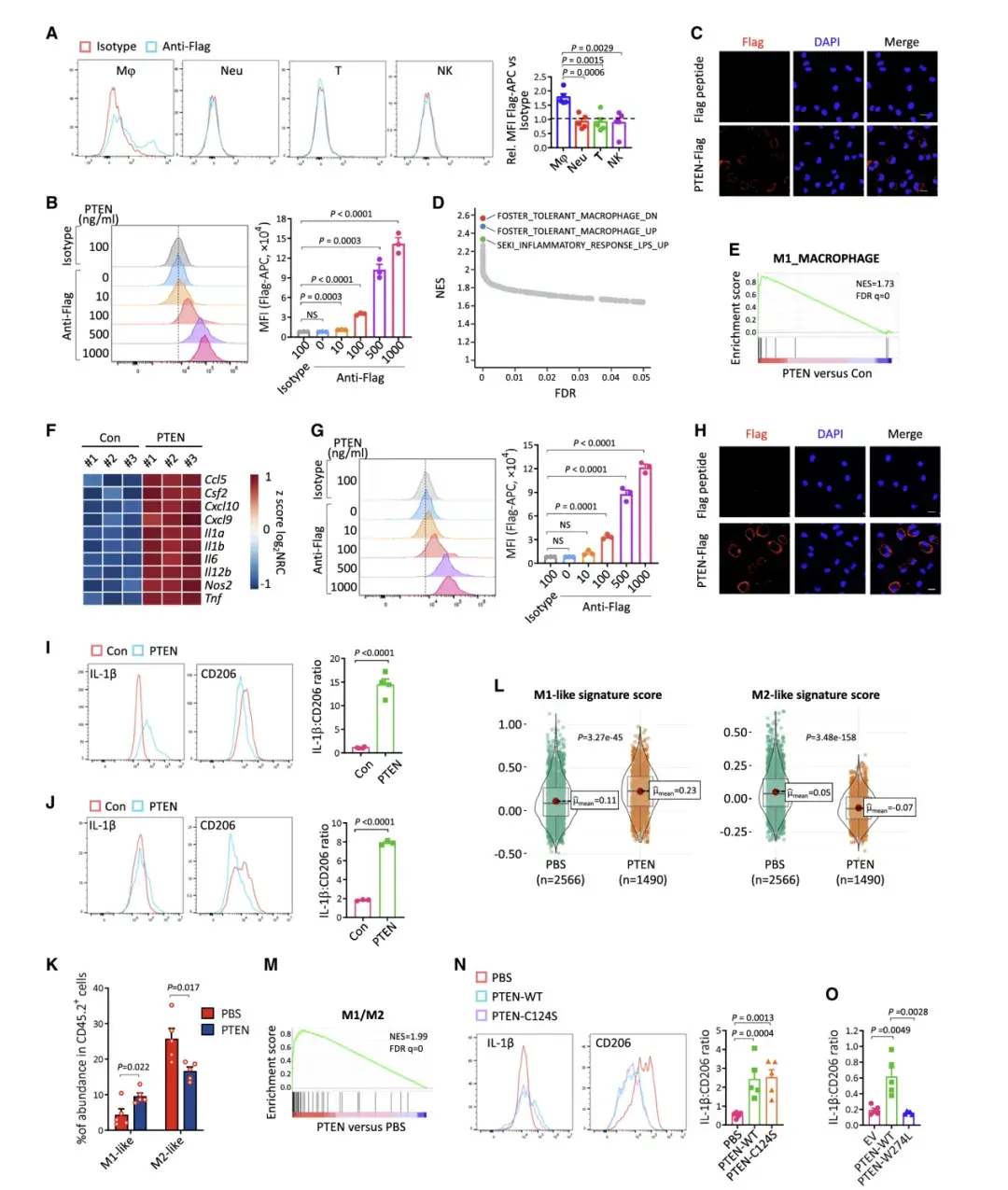
雖然 PTEN 治療廣泛改變了 BMDM 的轉錄組,但基因集富集分析 (GSEA) 表明,前三個上調通路都與脂多糖 (LPS) 刺激的信號傳導有關。
LPS 是一種常規刺激物,可誘導 BMDM 的促炎(M1 樣)極化, 這促使作者推測PTEN可能誘導BMDM發生M1樣極化。而GSEA和M1標志物表達的增加支持了這一觀點。這些結果表明,細胞外 PTEN 治療使BMDM 極化為 M1 樣表型。
TME中的巨噬細胞被稱為腫瘤相關巨噬細胞 (TAM),它們與免疫抑制性 M2 極化巨噬細胞非常相似。為了研究PTEN對TAM的影響,研究者先用IL-4和IL-13處理BMDM誘導M2樣極化。PTEN治療增加了M1標志物的表達,降低了M2標志物的表達,并顯著提高了IL-1b與CD206的表達比。這些結果表明,PTEN能將M2樣巨噬細胞轉化為M1樣表型。
先前CyTOF數據顯示 PTEN 治療的腫瘤具有明顯較少的 M2 樣巨噬細胞,卻有更多的 M1 樣巨噬細胞。另外,作者利用scRNA-seq 數據在TAM中計算了單細胞水平上基于 M1 和 M2 基因特征的分數,PTEN治療的腫瘤中分離的巨噬細胞顯示出明顯更高的 M1 基因特征分數,且M2基因特征分數低得多。RNA測序數據的GSEA分析也顯示PTEN處理的腫瘤發生了從M2到M1的轉化。流式細胞術分析表明PTEN治療增加了巨噬細胞中IL-1b與CD206的表達比率,且這種作用不依賴于PTEN的磷酸酶活性。PTEN-WT在腫瘤中的重新表達增加了IL-1b與CD206的表達比率,而PTEN-W274L的重新表達則顯示出大部分受損的作用。這些結果表明PTEN治療使TAM極化偏向M1樣表型。
04、PTEN 依賴巨噬細胞增強抗腫瘤免疫,抑制腫瘤生長
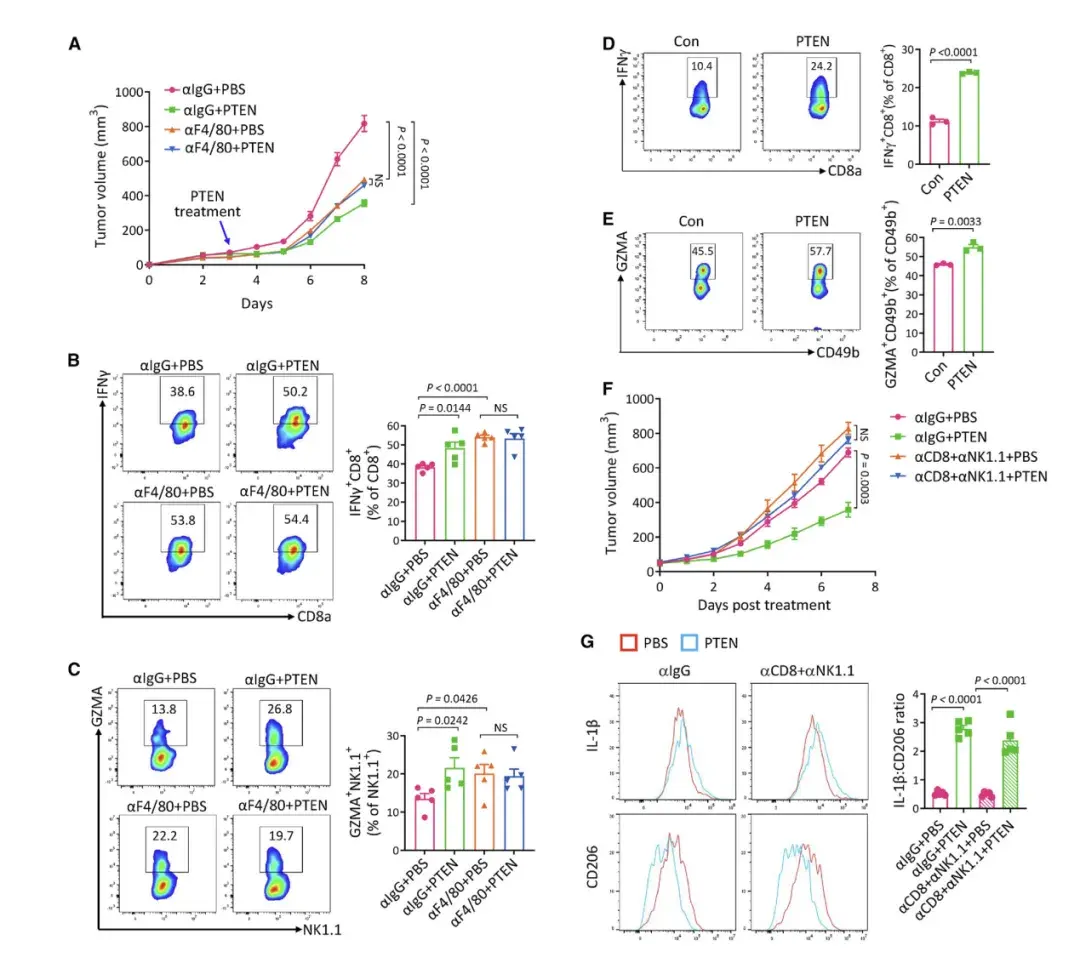
作者發現在缺乏PTEN的腫瘤中不再觀察到PTEN治療的抑癌作用,同時也不再促進CD8+ T和NK細胞的活化。此外,巨噬細胞的耗盡消除了PTEN 治療在促進 CD8+T和NK細胞活化方面的作用,這表明 TAM 是介導PTEN治療激活 CD8+T和NK細胞所必需的。這一觀點在體外得到了進一步證實。所有這些結果表明巨噬細胞極化是PTEN治療下CD8+T和 NK細胞激活的上游事件,它們的順序發生介導細胞外PTEN的腫瘤抑制作用。作者通過CellChat軟件分析scRNA-seq數據,發現SPP1-CD44軸在TAM抑制T細胞中起關鍵作用,并且PTEN處理顯著影響TAM和T細胞之間的相互作用。盡管PTEN治療增加了中性粒細胞的數量,但它們對于PTEN介導的腫瘤抑制并非必需。中性粒細胞的增加與巨噬細胞極化無直接關聯。因此,PTEN治療后中性粒細胞增加的機制和作用需要進一步研究。
05、PTEN 與巨噬細胞上的 PLXDC2 結合發揮抗腫瘤作用
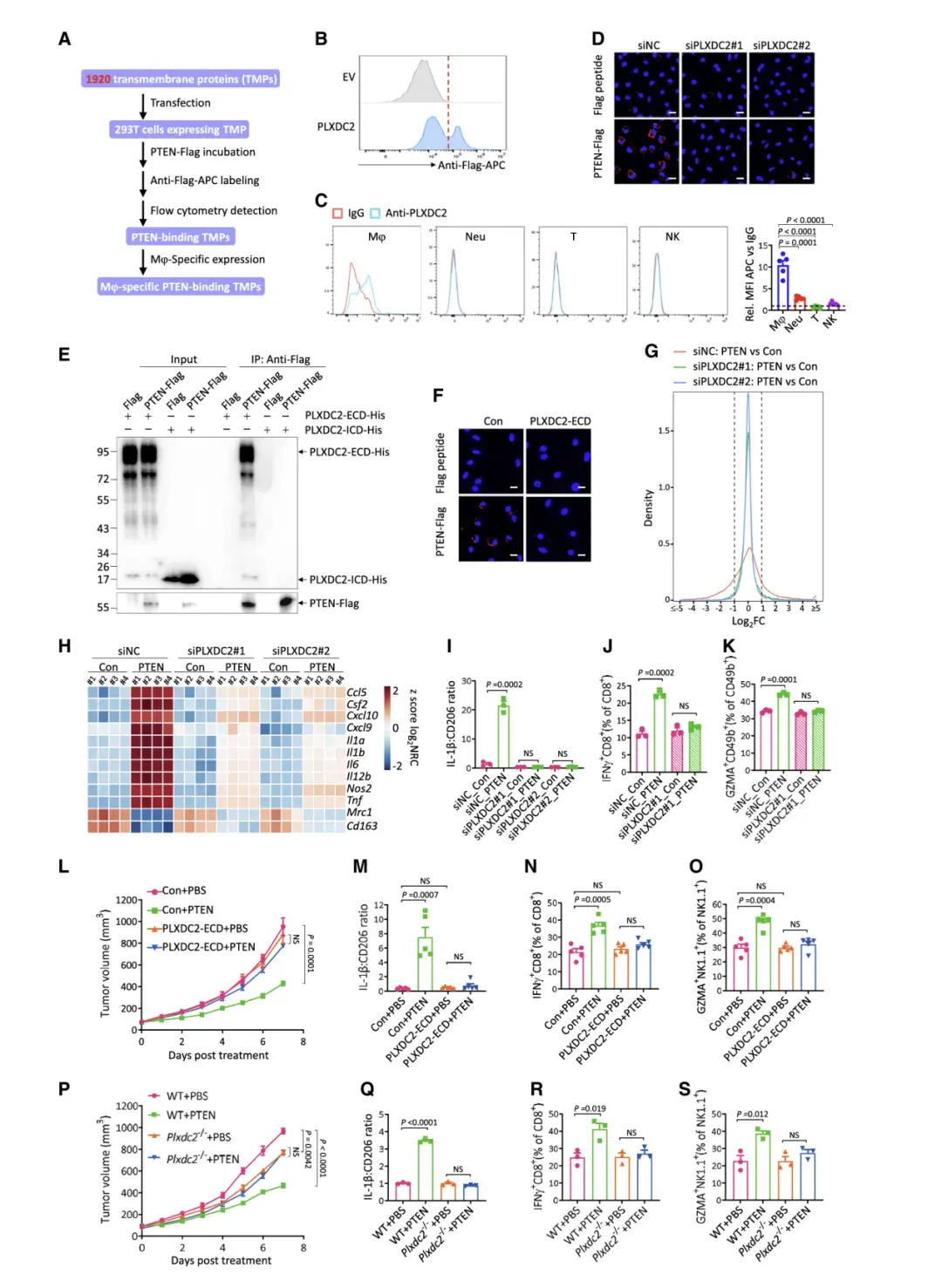
由于PTEN未進入巨噬細胞表面,而是優先與其結合,因此作者推測巨噬細胞存在特異性PTEN受體。為了識別該受體,作者通過數據庫篩選出了PLXDC2。PLXDC2 特異性地表達在巨噬細胞的表面,但不表達在中性粒細胞、T細胞或NK細胞的表面,敲低PLXDC2在很大程度上阻止了 PTEN 與M2極化的BMDM結合。
最后,作者發現PTEN在體外直接與PLXDC2的細胞外結構域 (ECD) 相互作用,但不與細胞內結構域 (ICD) 相互作用,并且不與細胞中缺乏ECD的PLXDC2相互作用。將過度純化的PLXDC2-ECD 添加到培養基中會消除PTEN與M2極化的BMDM表面的結合。這些結果都表明 PLXDC2 是一種介導 PTEN 與巨噬細胞表面結合的 PTEN 受體。
體外實驗顯示PTEN直接與PLXDC2的細胞外結構域相互作用。PLXDC2敲低顯著改變了PTEN處理下M2極化BMDM的轉錄組,逆轉了PTEN處理導致的基因變化。PLXDC2缺陷的BMDMs無法介導體外PTEN激活CD8+T和NK細胞的作用。使用PLXDC2敲低的巨噬細胞與B16-F10DPTEN細胞共植入小鼠,發現PTEN的腫瘤抑制作用在PLXDC2敲低組中顯著減弱。骨髓特異性PLXDC2耗盡小鼠實驗也顯示,PTEN處理對這些小鼠產生的腫瘤無抑制效果。這些結果表明,細胞外 PTEN 依賴于其與巨噬細胞表面的 PLXDC2 相互作用來激活抗腫瘤免疫并抑制腫瘤生長。
06、PTEN-PLXDC2 參與激活 JAK2-STAT1 信號以重新編程巨噬細胞
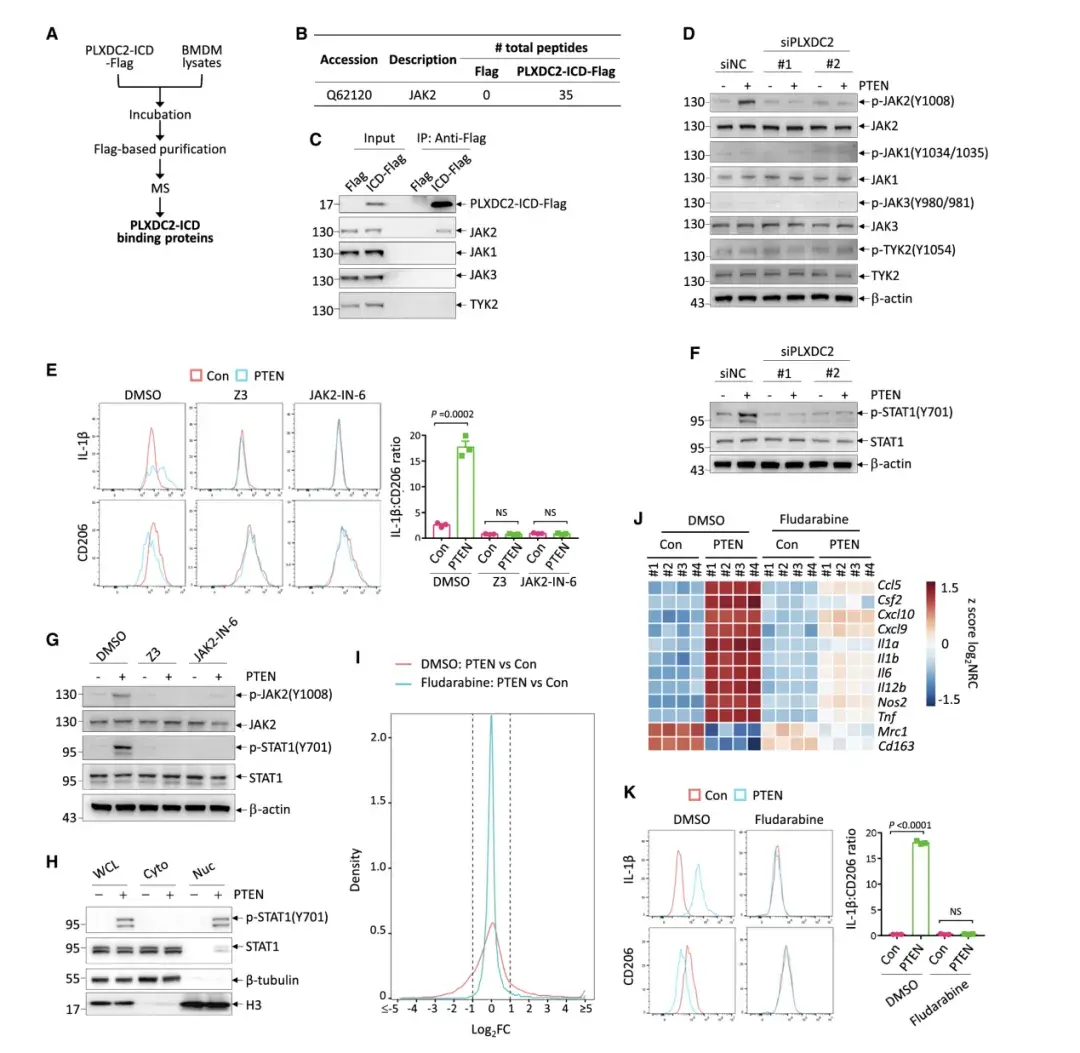
PLXDC2的下游信號傳導在很大程度上是未知的。作者發現在所有已確定與PLXDC2-ICD相互作用的蛋白質中,Janus 激酶 2 (JAK2) 表現出最強的強度。作者驗證了PLXDC2-ICD與 BMDM中的 JAK2而非其他家族成員的選擇性結合以及全長PLXDC2與293T細胞中異位表達的JAK2之間的相互作用。
試驗驗證了PLXDC2-ICD與JAK2的選擇性結合,并且PTEN處理增加了M2極化巨噬細胞中JAK2的磷酸化,這一過程依賴于PLXDC2。使用JAK2特異性抑制劑發現,它們能阻斷PTEN在巨噬細胞極化中的作用,表明PLXDC2通過PTEN激活JAK2來調節巨噬細胞極化。
TEN處理以PLXDC2和JAK2依賴的方式特異性地增加了 STAT1 (Tyr701) 的磷酸化,但未增加其他 STAT的磷酸化,表明PTEN PLXDC2-JAK2軸激活了STAT1。STAT1在巨噬細胞的M1極化中起著既定的作用。事實上,STAT1 抑制表現出與PLXDC2 敲低類似的效果,可拮抗細胞外PTEN對巨噬細胞重編程的影響。總的來說,作者確定了JAK2-STAT1信號傳導負責介導巨噬細胞中PTEN-PLXDC2的下游信號傳導。
07、細胞外PTEN與PD-1阻斷協同作用以抑制腫瘤生長
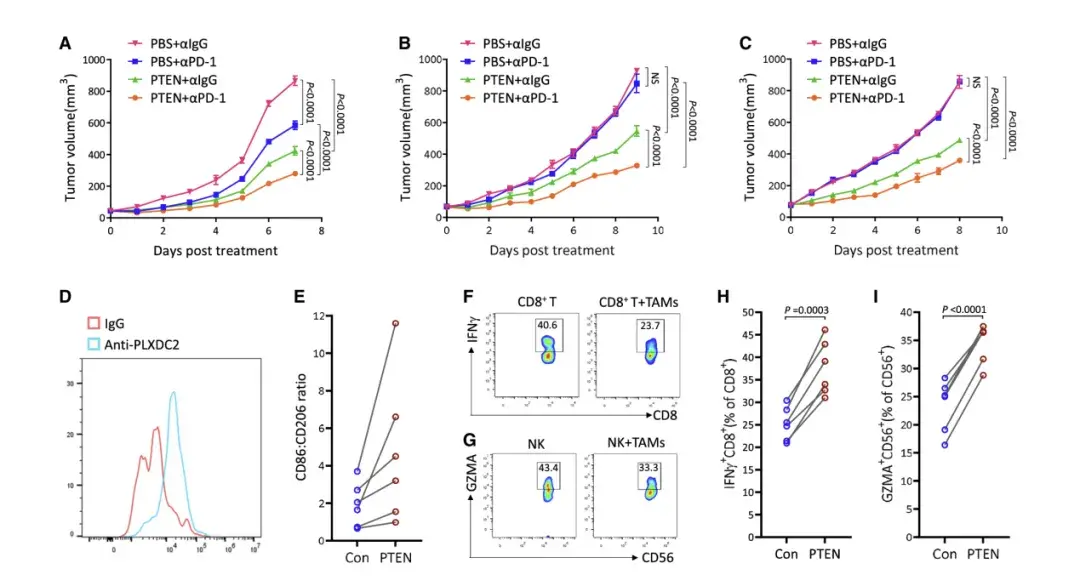
研究表明,M2 樣TAM對各種治療劑產生耐藥性,如PD-1阻斷。因此,作者測試了 PTEN 誘導的促腫瘤發生M2樣TAM 重編程為抗腫瘤 M1 樣表型是否會導致對免疫檢查點阻斷的反應增強。單用PTEN 或 PD-1 阻斷抗體治療都可縮小 B16-F10-DPTEN 腫瘤的大小,而聯合 PTEN/抗 PD-1 治療顯示出最顯著的效果。
PD-1阻斷不影響細胞外PTEN水平或TAM上PLXDC2表達,表明其作用獨立于PTEN-PLXDC2軸。在CT26結直腸癌和Lewis肺癌模型中,PD-1阻斷對腫瘤負荷無影響,但PTEN治療顯示出中等抗腫瘤作用,聯合治療則顯著減小了腫瘤大小。總體而言,PTEN治療增強了體內抗PD-1治療的效果。
為了解 PLXDC2在 TAM 中的臨床意義,作者創建了兩個基因特征來代表 PLXDC2 + 和 PLXDC2 TAM,并將它們應用于TCGA 數據庫中的大量 RNA-seq 數據集,使用 CIBERSORTx 工具來確定每個樣本中 PLXDC2 + 和 PLXDC2 TAM 的比例。分析顯示,PLXDC2 + TAM 比例較高的患者總生存率較低。在TCGA 數據庫的COAD、ESCA 和 LIHC 患者中也觀察到類似結果。這些發現與骨髓特異性 Plxdc2耗盡小鼠腫瘤生長延遲的結果一致。
從結腸直腸癌患者的腫瘤組織中分離出腫瘤浸潤性TAM,并確認其表達 PLXDC2。作者發現,所有患者的 TAM 在接受 PTEN 治療后,CD86 與 CD206 的表達比率均增加,表明 PTEN 誘導患者來源的 TAM 的 M2 極化至 M1 極化。作者還從每位患者的外周血中分離出 CD8+ T 和NK 細胞,并使用來自一名患者的樣本示例性地證實了自體 TAM 抑制了這些細胞。然后,每個患者的TAM經PTEN處理后與自體CD8+ T和NK細胞孵育。結果表明,與 PTEN 處理的 TAM 一起孵育的 CD8+ T 和 NK 細胞比與未用 PTEN 處理的 TAM 一起培養的 CD8+ T 和 NK 細胞表現出更高的活性,表明 PTEN 在減輕患者來源的 TAM 的免疫抑制作用方面發揮作用。因此,體外數據表明,患者活組織檢查的TAM對PTEN治療有反應。
派森諾目前已致力于單細胞測序多年,憑借其豐富的項目經驗、出色的服務能力和在疑難樣本解離方面獨特經驗,贏得了業界的廣泛贊譽。目前,派森諾生物單細胞轉錄組測序極速周期活動正在火熱進行中,詳情請聯系當地銷售。