

2017-07-19
SlARF10,番茄生長素響應因子,受Sl-miR160的轉錄調控。通過表達SlARF10(mSlARF10),發現其小葉葉片比較窄,葉片上有較大的氣孔但密度比較低。35S:mSlARF10-6植株和野生型WT相比, 其窄型離體葉片失水較多。研究發現了之前未識別到的一個葉片失水調控因素,以及在這個重要過程中生長素和ABA信號協調作用的互作網絡關系。miR160調控ARF10,來維持葉片中的水平衡,以保證正常的植株生長和環境適應性。
1、研究發現在35S:mSlARF10-6植株和野生型WT中,實際的失水量和計算的氣孔的失水量是不一致的,表明他們存在導水率差異。過表達mSlARF10可能會通過蒸騰作用增加了轉基因植株的葉片的失水量。35S:mSlARF10-6葉片的導水率要比WT中顯著提高(圖1b)。結果顯示mSlARF10過表達不僅改變葉片性狀,而且影響導水率。
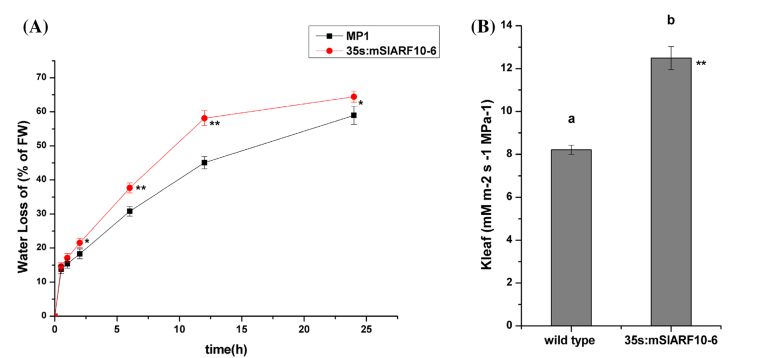
圖1,過表達mSlARF10增加了葉片失水。a:葉片失水率,b:葉片導水率
2、用ABA(脫落酸)和HgCl2做預處理,驗證在35S:mSlARF10中有較高的導水率,這和其有較大的氣孔和較高活性水通道蛋白相關(AQPs)。ABA處理的條件下,35S:mSlARF10-6有著較高的敏感性,氣孔會迅速關閉。在WT的0-24h的脫水處理中,內源性ABA的積累逐漸增加。ABA敏感性實驗檢測SlARF10是否和ABA信號傳導相關。在35S:mSlARF10-6和WT葉片中,ABA處理可能減少了失水(圖2)。ABA處理的35S:mSlARF10-6中葉片的失水很大程度的下降,將近20%。ABA處理的WT的葉片失水量在脫水處理6h后,比脫水處理6h前還低。35S:mSlARF10-6中失水速度比WT中要高,但是ABA處理后,在脫水處理6h前要比WT中要慢很多。這表明在早期的脫水處理中,ABA在35S:mSlARF10-6葉片中對失水的抑制作用要比WT中要大。
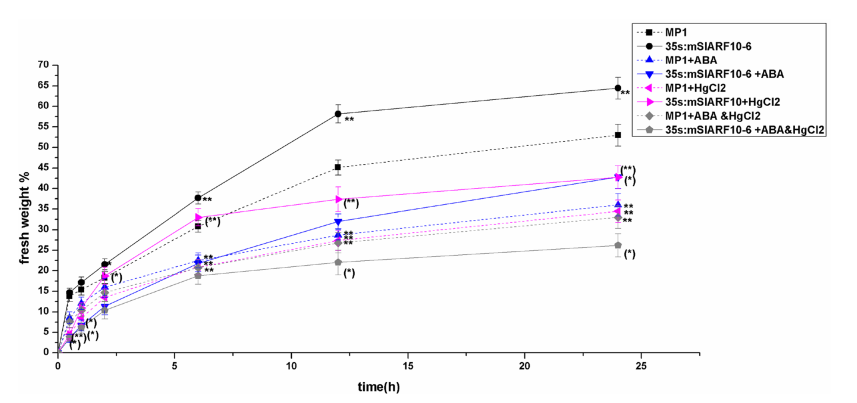
圖2,不同處理條件下,葉片的失水率
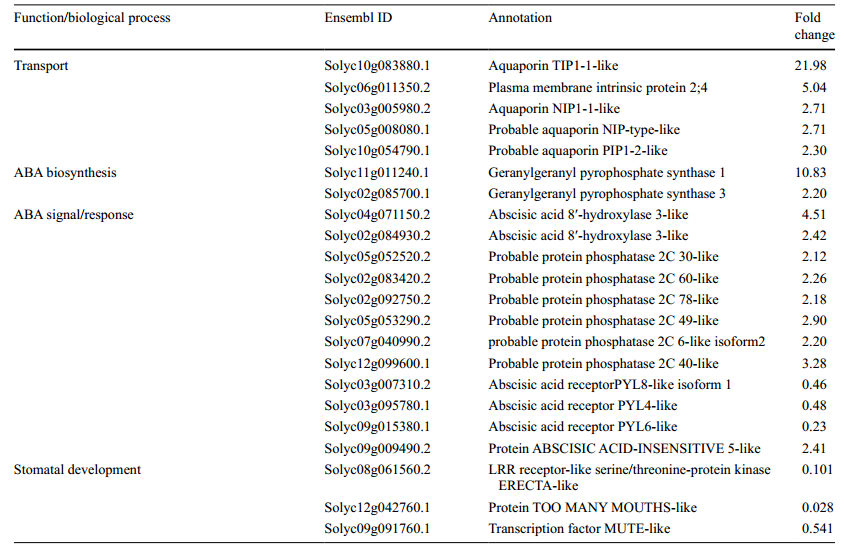
3、RNA測序表明5種AQP相關的基因,14種ABA合成/信號傳導基因和2種氣孔發育基因,在35S:mSlARF10-6植株中明顯被改變,這一結果被qRT-PCR驗證。用WT和35S:mSlARF10-6的15個葉片做RNA測序,檢測35S:mSlARF10-6葉片對整體ABA合成,信號傳導,和AQP基因表達的影響。結果發現在35S:mSlARF10-6樣本中發現5個顯著上調的QP基因,2個ABA合成和12個信號/響應顯著上調或下調的基因(表1)。
表1,葉片中差異性上調的AQPs,ABA和氣孔發育相關的基因
4、實驗發現,SlARF10通過調節氣孔開合度來緩和失水現象,增強了ABA合成/信號響應;也通過影響氣孔的形成和AQP的表達來影響水的運輸,二者一起協調控制番茄葉片的失水。
構建互作反應的網絡圖,說明miR160在ARF10在植物水壓環境中的調控作用中的精確作用(圖3)。該模式說明ABA-植物生長素相互作用對控制失水的影響。當生長素信號被激活,Aux/IAA被分解,ARF10被高水平轉錄。盡管ARF10可以通過ABA合成基因活躍的表達增強ABA含量,通過ABI5表達增強ABA信號,最后導致干旱是氣孔關閉。適當表達水平受miRNA160調控的ARF10,可能在葉片發育中維持葉片水平衡和適應水壓中有著重要的作用。
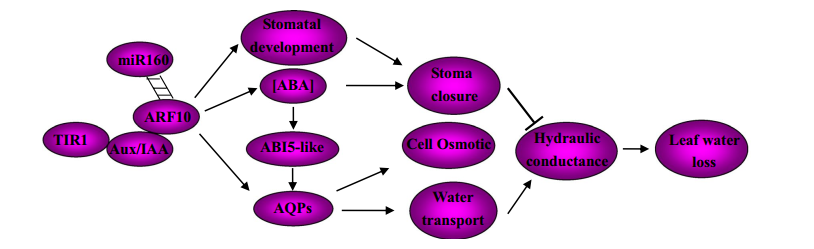
圖3,針對miR160調控的ARF10在對葉片失水的響應中起的作用提出了一種互作模式
結論:
1、 mSlARF10的過表達導致葉片變窄和更高的失水率
2、 mSlARF10的過表達通過影響ABA合成和信號傳導調控氣孔關閉
3、 mSlARF10的過表達誘導葉片表皮的AQP表達
4、 受miR160調控的SlARF10控制葉片中生長素和ABA對失水現象的響應


















