馬鈴薯膨大的含淀粉的塊莖、柑橘富含維生素C的果實(shí)、辣椒的口感辛辣的果實(shí)、園林植物多樣的花形、藥用植物具生物活性的次生代謝產(chǎn)物……高等植物豐富多樣的特化的性狀廣泛吸引了研究者的關(guān)注。目前,研究熱點(diǎn)主要集中于具有生產(chǎn)效益和經(jīng)濟(jì)價(jià)值的性狀上,研究方向涉及果實(shí)發(fā)育成熟、特異代謝產(chǎn)物合成、農(nóng)藝性狀(產(chǎn)量、品質(zhì))、園藝性狀(花形、花期)、植株病蟲(chóng)抗性等方面。通過(guò)基因de novo測(cè)序可以一次鑒定到目標(biāo)性狀的多個(gè)相關(guān)基因,發(fā)現(xiàn)新的基因位點(diǎn),揭示性狀形成和調(diào)控的遺傳機(jī)制,為進(jìn)一步的驗(yàn)證和應(yīng)用提供指導(dǎo)。
性狀相關(guān)基因鑒定分析的實(shí)驗(yàn)設(shè)計(jì)主要包括種間比較和種內(nèi)比較兩個(gè)途徑。種間比較常通過(guò)遺傳和表型差異較大的品種間比較、同屬或同科的近緣有參物種間比較,或以模式植物擬南芥為參考進(jìn)行比較,鑒定并比較目標(biāo)性狀相關(guān)的同源基因,比較內(nèi)容涉及基因表達(dá)情況和拷貝數(shù)等。種內(nèi)比較通常考察相關(guān)基因的表達(dá)情況,通過(guò)在根、莖、葉、花、果等不同組織部位取樣,或在不同生長(zhǎng)發(fā)育階段取樣,來(lái)比較目標(biāo)基因的表達(dá)差異并分析其與特異性狀的關(guān)系。基因表達(dá)模式的檢測(cè)常采用轉(zhuǎn)錄組測(cè)序或表達(dá)譜測(cè)序,或進(jìn)行拷貝數(shù)檢測(cè)等。
對(duì)于實(shí)驗(yàn)結(jié)果的討論可結(jié)合研究物種的特點(diǎn),綜合進(jìn)化分析內(nèi)容,如基因家族的擴(kuò)張/收縮、全基因組倍增事件、多倍化事件等驅(qū)動(dòng)高等植物進(jìn)化的事件或馴化改良施加的選擇壓力,提出目標(biāo)基因可能的形成原因,闡釋目標(biāo)性狀產(chǎn)生的遺傳和分子機(jī)制,并規(guī)劃進(jìn)一步的驗(yàn)證工作。通過(guò)以下文獻(xiàn)案例,我們可對(duì)這方面的研究思路窺見(jiàn)端倪。
1. 辣椒果實(shí)發(fā)育及辣椒素合成相關(guān)基因
這是2014年P(guān)NAS的一篇文章,通過(guò)對(duì)同屬茄科但分屬不同果實(shí)發(fā)育模式的辣椒和番茄的比較,鑒定了果實(shí)發(fā)育相關(guān)基因,通過(guò)對(duì)辛辣和不辣的辣椒品種的比較及轉(zhuǎn)錄組測(cè)序,鑒定了辣度相關(guān)基因并分析了其表達(dá)模式。
果實(shí)的成熟過(guò)程顯著影響果實(shí)品質(zhì)和保存期限,在躍變型果實(shí)(番茄)和非躍變型果實(shí)(辣椒)間差異顯著。非躍變型果實(shí)軟化過(guò)程更為緩慢且對(duì)乙烯無(wú)響應(yīng)。比較番茄和辣椒在果實(shí)成熟期的表達(dá)譜,發(fā)現(xiàn)番茄有2281個(gè)特異基因,而辣椒有1440個(gè)特異基因,這兩個(gè)物種的特異基因均涉及細(xì)胞壁重構(gòu)、激素信號(hào)傳導(dǎo)和代謝、碳水化合物代謝、蛋白質(zhì)降解以及非生物逆境響應(yīng)等。
但兩者的差異在于以下4點(diǎn):1)涉及乙烯生物合成的基因數(shù)目在辣椒中更低,辣椒的8個(gè)編碼1-氨基環(huán)丙烷-1-羧酸鹽合酶(乙烯生產(chǎn)的關(guān)鍵酶)的基因在果實(shí)成熟過(guò)程中無(wú)一上調(diào),與辣椒較低的乙烯合成一致,而番茄中的2個(gè)該基因被強(qiáng)烈誘導(dǎo);2)在辣椒中與乙烯信號(hào)和茉莉酸信號(hào)傳導(dǎo)相關(guān)的差異表達(dá)基因的數(shù)目更低;3)與生長(zhǎng)素和脫落酸相關(guān)的差異表達(dá)基因數(shù)目更大,包括與非生物逆境相關(guān)的基因,這與草莓(非躍變型果實(shí))成熟過(guò)程中脫落酸的積累一致;4)葉片衰老的負(fù)調(diào)控因子WRKY70和ZAT10在辣椒中受到更強(qiáng)烈的誘導(dǎo),表明這些轉(zhuǎn)錄因子的誘導(dǎo)可能對(duì)于辣椒的長(zhǎng)保存期起到重要作用;5)9個(gè)番茄XTH(木葡聚糖內(nèi)源轉(zhuǎn)糖基酶/水解酶)基因中的15個(gè)在果實(shí)成熟過(guò)程中呈現(xiàn)差異表達(dá),而辣椒的25個(gè)XTH基因中只有6個(gè)差異表達(dá),表明XTH活性降低導(dǎo)致辣椒果實(shí)成熟過(guò)程中軟化減弱。
辣椒素的積累,主要包括辣椒素和二氫辣椒素,為辣椒屬植物獨(dú)有且導(dǎo)致了果實(shí)的辛辣度(辣椒素合成通路見(jiàn)圖1)。基于對(duì)辣椒辣度的前期研究,在辣椒、番茄、馬鈴薯和擬南芥中,鑒定了51個(gè)涉及辣椒素合成的基因家族及其同源基因。進(jìn)化分析顯示,與其他3個(gè)物種相比,辣椒有獨(dú)立的、特有的13個(gè)基因家族的重復(fù)(如ACLd、AT3、β-CT、C3H、CAD、CCR、Kas I和PAL)。基因重復(fù)中的序列差異可能導(dǎo)致功能差異或新功能的產(chǎn)生,促進(jìn)特異性的辣椒素生物合成的進(jìn)化。
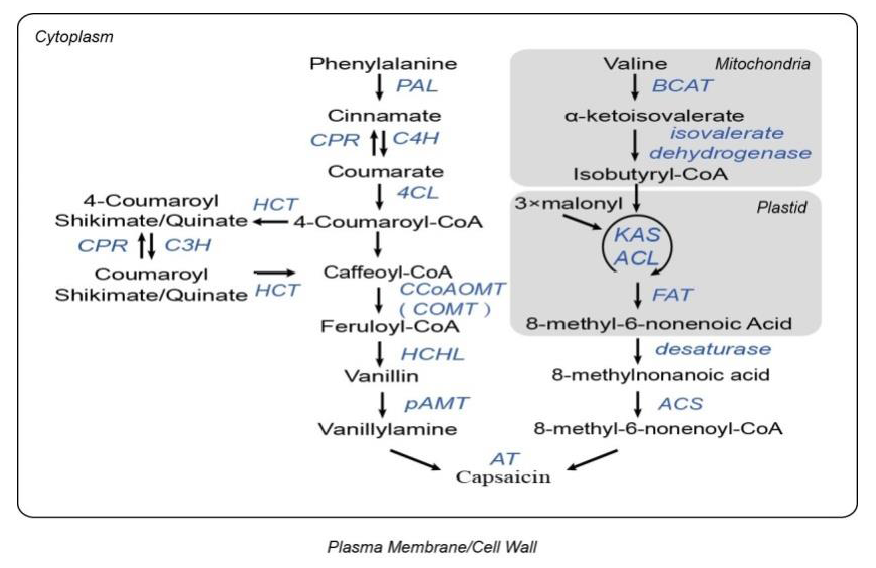
圖1. 辣椒素生物合成通路,來(lái)源:文獻(xiàn)補(bǔ)充資料
以AT3為例,在辣椒中鑒定了3個(gè)At3(Pun1)的串聯(lián)拷貝,編碼假定的酰基轉(zhuǎn)移酶且在某些辣椒屬植物中對(duì)辣度起調(diào)控作用。野生和栽培辣椒中AT3-D1和AT3-D2均在保守的DFGWGKP結(jié)構(gòu)域有氨基酸替換。對(duì)AT3-D1的分析表明(圖2),在非辛辣基因型中,pun1等位基因(C位點(diǎn))有2724/2930bp的缺失,橫跨假定的啟動(dòng)子區(qū)和第一外顯子。在辛辣的辣椒品種中,AT3-D1和AT3-D2(圖3)同樣鑒定到了短InDel和單堿基非同義替換。
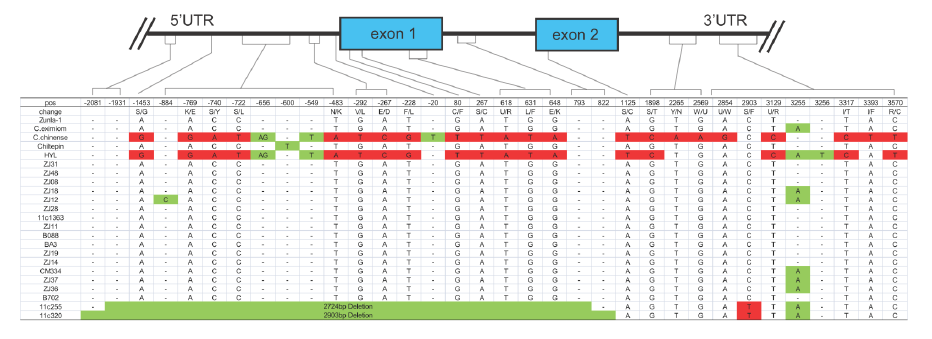
圖2. AT3-D1基因結(jié)構(gòu),來(lái)源:文獻(xiàn)補(bǔ)充資料
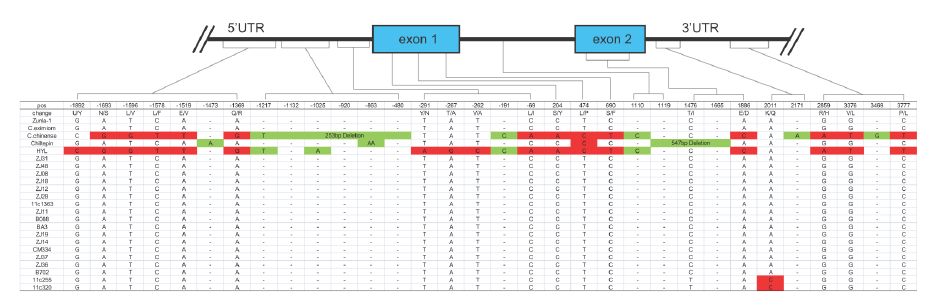
圖3. AT3-D2基因結(jié)構(gòu),來(lái)源:文獻(xiàn)補(bǔ)充資料
辣椒素生物合成相關(guān)基因的組織特異性和發(fā)育相關(guān)表達(dá)模式檢測(cè)發(fā)現(xiàn)(圖4),多數(shù)基因家族,除ACL-D4和ACL-D5外,伴隨辣椒素積累呈現(xiàn)組織和發(fā)育階段特異性的表達(dá)模式。然而CCoAOMT-D9、AT3-D1和AT3-D2只在辣椒素合成的果實(shí)發(fā)育階段顯著表達(dá)。上述基因在5個(gè)非辛辣種的表達(dá)模式顯示,AT3-D1檢測(cè)不到或微量,表達(dá)缺失可能由于pun1等位基因的大片段缺失,使其成為非辛辣辣椒中的假基因。有趣的是,在非辛辣種中,AT3-D2的表達(dá)或能保持微量的辣椒素和二氫辣椒素。推測(cè)AT3-D1和AT3-D2在C位點(diǎn)的劑量補(bǔ)償效應(yīng)形成了辣椒中辣度的差異。
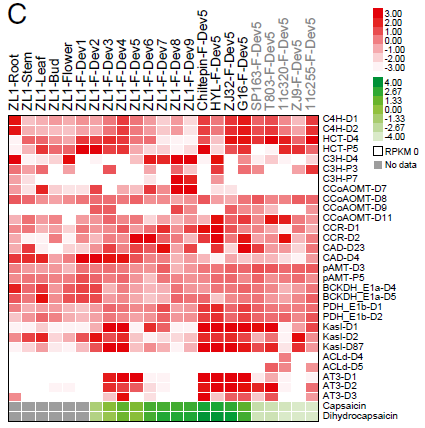
圖4. 辣椒素生物合成相關(guān)基因表達(dá)模式,來(lái)源:文獻(xiàn)原文
2. 棉纖維形成和伸長(zhǎng)相關(guān)基因分析
這篇2012年的Nature Genetics文章通過(guò)對(duì)開(kāi)花后3天(DPA)胚珠的轉(zhuǎn)錄組測(cè)序,在無(wú)纖維的雷蒙德氏棉和有纖維的陸地棉之間發(fā)現(xiàn)了關(guān)鍵成纖維基因的轉(zhuǎn)錄本的定量差異,包括Sus、KCS、ACO、MYB和bHLH基因。
鑒別到的4種蔗糖合酶(Sus)基因中,3種(SusB、Sus1和SusD)在陸地棉中表達(dá)量大幅高于雷蒙德氏棉。(圖5)
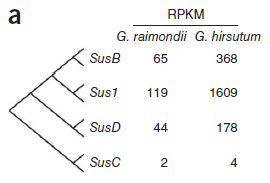
圖5. Sus基因表達(dá)量差異,圖片來(lái)源:文獻(xiàn)原文
若干種3-酮乙基-輔酶A合酶(KCS)基因,包括KCS2、KCS13和KCS6只在陸地棉中表達(dá),而兩種棉中都存在中等表達(dá)量的KCS7基因。表明高表達(dá)水平的Sus和KCS基因家族對(duì)于纖維細(xì)胞的形成和伸長(zhǎng)可能確為必需的。(圖6)
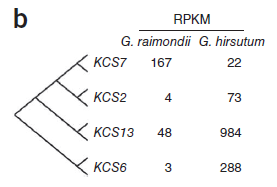
圖6. KCS基因表達(dá)量差異,圖片來(lái)源:文獻(xiàn)原文
相對(duì)的,編碼1-氨基環(huán)丙烷-1-羥酸氧化酶(ACO)的轉(zhuǎn)錄本在雷蒙德氏棉存在極端高含量,表明乙烯在纖維細(xì)胞發(fā)育早期有重要作用。(圖7)
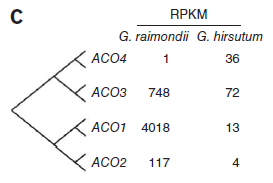
圖7. ACO基因表達(dá)量差異,圖片來(lái)源:文獻(xiàn)原文
此前研究提出假設(shè),即棉纖維在形態(tài)和起源上與植物表皮毛相似,表皮毛為多種植物組織中存在的毛狀表皮細(xì)胞,常見(jiàn)于葉片和莖表面。假設(shè)提出,在擬南芥表皮毛發(fā)育中起重要作用的轉(zhuǎn)錄因子可能與棉纖維形成相關(guān)。在擬南芥中,MYB和bHLH類(lèi)轉(zhuǎn)錄因子與TTG1形成復(fù)合物共同起作用調(diào)控表皮細(xì)胞發(fā)育。總計(jì)2706個(gè)轉(zhuǎn)錄因子,包括208個(gè)bHLH和219個(gè)MYB基因在雷蒙德氏棉基因組中得到鑒定。大量的MYB和bHLH基因在陸地棉胚珠中顯著表達(dá),而雷蒙德氏棉胚珠中只有殘量表達(dá),表明這些基因的部分可能為纖維早期發(fā)育所必需。(圖8)
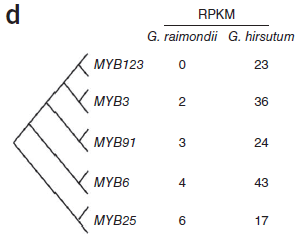
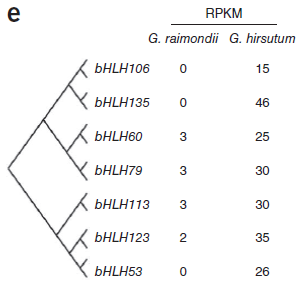
圖8. MYB和bHLH基因表達(dá)量差異,圖片來(lái)源:文獻(xiàn)原文
這篇文章同樣對(duì)棉子酚生物合成基因進(jìn)行了鑒定和進(jìn)化分析。棉會(huì)產(chǎn)生一種獨(dú)有的萜類(lèi),包括脫氧半棉酚、半棉酚、棉子酚、半棉酚酮、殺實(shí)夜蛾素。棉植株在色素腺體中積累棉子酚及相關(guān)倍半萜類(lèi)來(lái)防御病蟲(chóng)害。大部分棉倍半萜類(lèi)都有一個(gè)共同前體衍生出來(lái),(+)-δ-杜松萜烯,由(+)-δ-杜松萜烯合酶(CDN)通過(guò)法尼基二磷酸的環(huán)化作用合成,是棉子酚生物合成的第一個(gè)關(guān)鍵步驟。以前,CDN-A和CDN-C均被報(bào)道編碼CDN酶活性。利用雷蒙德氏棉和其他8種有參物種的進(jìn)化分析表明,除了水稻,萜環(huán)化酶基因家族為多種植物所共有。然而,只有雷蒙德氏棉和可可(同屬錦葵目Malvales)擁有具生化功能的CDN1基因家族。似乎棉子酚合成能力與古六倍體和全基因組倍增事件均相關(guān)相關(guān)。在近緣分支的番木瓜和毛果楊中沒(méi)有發(fā)現(xiàn)CDN1的同源基因,表明棉子酚的產(chǎn)生是在這些物種分離之后出現(xiàn)的。DDtYD和DDVAE結(jié)構(gòu)域是棉子酚生物合成的關(guān)鍵因素,其他植物萜環(huán)化酶基因不編碼含DDVAE結(jié)構(gòu)域的蛋白因此不屬于CDN同源基因。
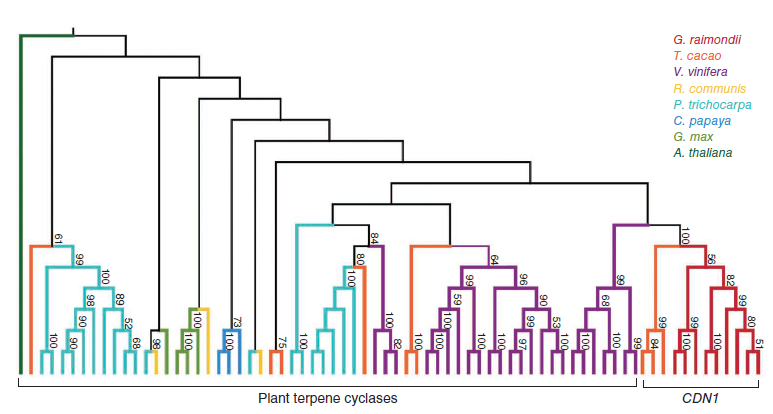
圖9. 棉子酚CDN1基因家族進(jìn)化分析,圖片來(lái)源:論文原文
3. 木槿的花期和抗病基因
2017年DNA Research的一篇文章做了木槿的de novo測(cè)序并研究了花期調(diào)控和植株抗性相關(guān)基因。不同植物花發(fā)育的的遺傳和分子機(jī)制是高度保守的,包含4種主要的開(kāi)花通路(光周期、自主調(diào)控、春化作用和赤霉素)。光周期通路中主要的開(kāi)花信號(hào)受到FLOWERING LOCUS T (FT)的調(diào)控,而春化作用通路是通過(guò)暴露于特定刺激物后FT抑制劑的去除起作用。木槿是長(zhǎng)日照開(kāi)花植物,花期長(zhǎng)且每天開(kāi)花20~30朵不等。而一朵花只開(kāi)放一天。為了揭示調(diào)控這些表型的遺傳機(jī)制,研究了無(wú)油樟、擬南芥、可可、雷蒙德氏棉和木槿中4個(gè)開(kāi)花通路涉及的基因,且研究了花、子房、根和葉等不同木槿組織中的基因表達(dá)模式。
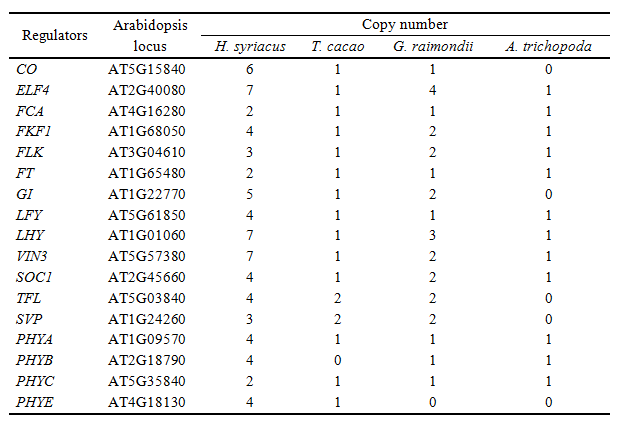
花期基因的進(jìn)化分析識(shí)別到木槿特異性的分支。由于花期經(jīng)常取決于基因拷貝數(shù),使用擬南芥的基因?yàn)閰⒖迹缍ú煌参锘蚪M中拷貝數(shù)變化,發(fā)現(xiàn)木槿的拷貝數(shù)為其余4者的2~7倍。本研究涉及的花期調(diào)控基因中,涉及晝夜節(jié)律調(diào)節(jié)(CO,ELF4, FKF1, GI, LHY, PHYs)和花芽形成(FCA, FLK, FT,LFY, VIN3, SOC1, TFL, SVP)基因數(shù)在木槿中顯著提高。此外,調(diào)控光敏色素A信號(hào)的FAR1家族基因的拷貝數(shù)在木槿中更高。穗狀花序的植物,如大麥、水稻、小麥,同樣包含FAR1基因的高拷貝數(shù),因此,FAR1的高拷貝數(shù)或可同樣影響木槿的表型。(表1)
表1. 花期基因拷貝數(shù)比較,圖片來(lái)源:論文原文
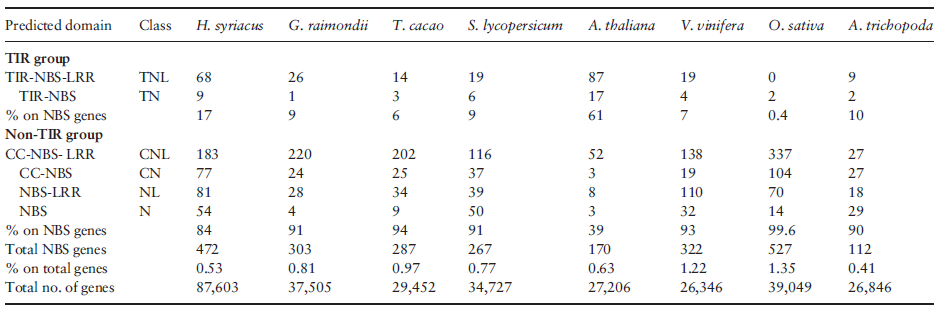
多數(shù)抗病基因家族(R)編碼含NBS和富含亮氨酸重復(fù)序列(LRR)的胞內(nèi)蛋白。編碼NBS的R基因家族是木槿最大的基因家族之一,含472個(gè)基因,約為無(wú)油樟和擬南芥的3倍。這些基因基于Toll/白細(xì)胞介素受體(TIR)結(jié)構(gòu)域的差異分為2個(gè)分支。相比番茄、雷蒙德氏棉和可可,木槿中TIR基因顯著過(guò)表達(dá)。錦葵目植物(木槿、可可、雷蒙德氏棉)中多于70%的編碼NBS的基因在26個(gè)亞類(lèi)中共享,表明多數(shù)R基因由共同祖先衍生而來(lái)。另外,木槿中來(lái)自4個(gè)亞類(lèi)的125個(gè)NBS編碼基因約比其他錦葵科植物擴(kuò)增了5倍,而來(lái)自7個(gè)亞類(lèi)的18個(gè)NBS編碼基因?yàn)槟鹃泉?dú)有。木槿基因組中TIR和RPW8編碼基因亞類(lèi)顯示出廣泛的擴(kuò)張,經(jīng)歷了不均等復(fù)制事件,顯示出植物基因組中高度多樣性。木槿基因組中不同的R基因組成提示成簇的R基因的擴(kuò)張和多樣性可能涉及種系特異性的基因倍增事件,最終導(dǎo)致近緣種的趨異進(jìn)化。這些結(jié)果為多年生植物中花期和抗病基因的進(jìn)一步比較分析提供了初步的信息支持。(表2)
表2. NBS-LRR基因家族數(shù)目比較
參考文獻(xiàn)
[1] Qin C, Yu C, Shen Y, et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization[J]. Proceedings of the National Academy of Science, 2014, 111(14):5135-40.
[2] Wang K, Wang Z, Li F, et al. The draft genome of a diploid cotton Gossypium raimondii.[J]. Nature Genetics, 2012, 44(10):1098.
[3] Kim Y M, Kim S, Koo N, et al. Genome analysis of Hibiscus syriacus provides insights of polyploidization and indeterminate flowering in woody plants[J]. Dna Research An International Journal for Rapid Publication of Reports on Genes & Genomes, 2017, 24(1):71-80.




















