M6A依賴的母體mRNA清除促進斑馬魚的母源-合子過渡
2017-09-27
研究背景
m6A(N6-methyladenosine,6-甲基腺嘌呤)是真核生物mRNA內(nèi)部序列中最常見的一種甲基化修飾,同時可以功能性的調(diào)節(jié)真核生物的轉(zhuǎn)錄組從而影響mRNA的剪接、出核、定位、翻譯和穩(wěn)定。m6A RNA出現(xiàn)在多種重要的細胞生命過程中,意味著mRNA甲基化參與了多種生物學(xué)過程,如干細胞分化、生物節(jié)律等。最近新發(fā)現(xiàn)的m6A 識別蛋白YTHDF2可以對 m6A 去甲基化。
哺乳動物早期胚胎發(fā)育過程中,胚胎中許多蛋白的產(chǎn)生依賴于來自母源的mRNA調(diào)控的基因表達,但是在經(jīng)歷母體-合子過渡(maternal-to-zygotic transition)期間后,胚胎將激活自身的基因組從而產(chǎn)生變化,此后大量來自母源的mRNA將被迅速清除。斑馬魚ythdf2在早期胚胎中普遍表達。本研究通過在斑馬魚發(fā)育早期階段使ythdf2喪失功能來檢測m6A 依賴性RNA是否減少。
研究結(jié)果
實驗表明,純合的第一代ythdf2-/-斑馬魚突變體沒有產(chǎn)生表型,但將純合的第一代ythdf2-/-斑馬魚進行雜交時發(fā)現(xiàn)大約有超過70%的后代無法發(fā)育通過第一個細胞期,然后進一步發(fā)現(xiàn)這種致死可能是由于純合ythdf2-/-雄性斑馬魚的精子缺陷導(dǎo)致。
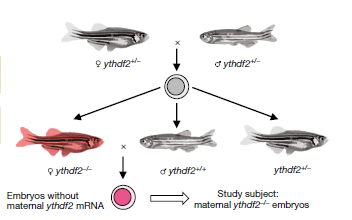
圖1.斑馬魚早期胚胎樣本構(gòu)建
利用m6A-seq發(fā)現(xiàn),40% 的基因轉(zhuǎn)錄本存在m6A位點,超過36%的母體基因具有m6A甲基化位點。受精后4h之前,甲基化基因比非甲基化基因豐度更高且主要富集在磷代謝和細胞周期調(diào)控功能上。
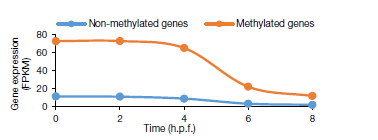
圖2.未甲基化基因與甲基化基因比對
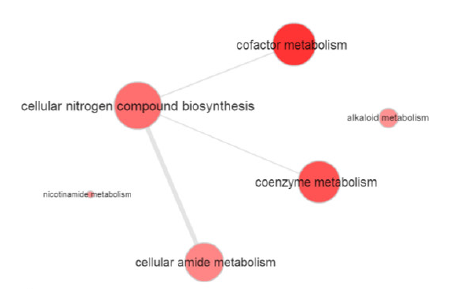
圖3.甲基化基因GO富集分析
進一步研究發(fā)現(xiàn),與野生型相比,受精后4h時,ythdf2?/?胚胎中mRNA的m6A甲基化修飾提高了50%以上,證明Ythdf2會影響母體mRNA的甲基化。而且它們的清除會因為YTHDF2的缺失而受阻。對受精后4h時間點下母體ythdf2?/?樣本進行測序分析,共發(fā)現(xiàn)3292個基因高表達并存在甲基化,其中超過60%為母體基因。經(jīng)過篩選,其中135個基因為共有基因,這些共有基因主要富集在磷代謝、細胞周期過程和繁殖過程中。
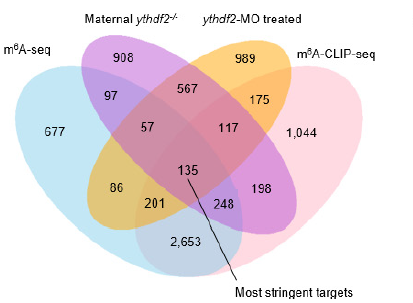
圖4.受精后4h時間點利用不同測序技術(shù)發(fā)現(xiàn)的m6A甲基化修飾轉(zhuǎn)錄本
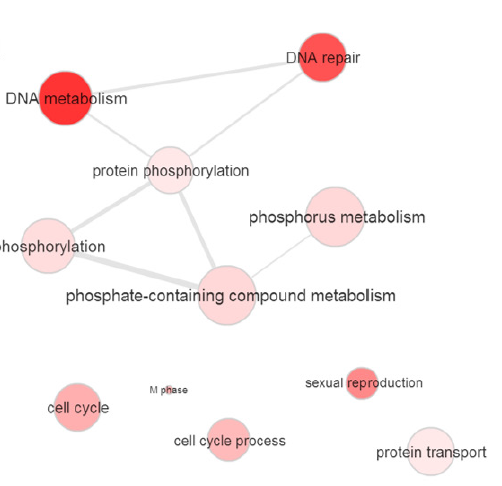
圖5.共有基因GO富集分析

圖6.RNA甲基化motif
研究結(jié)論:
本研究表明,m6A甲基化修飾在早期胚胎發(fā)育中具有非常關(guān)鍵的作用。超過三分之一的斑馬魚母本mRNA可以通過N6甲基腺苷(m6A)進行修飾調(diào)控,而且這些母本mRNA清除過程也是通過m6A結(jié)合蛋白Ythdf2完成的。在斑馬魚胚胎中去除Ythdf2,會減緩了m6A修飾母本mRNA的衰變,并且阻止合子基因組激活。胚胎無法及時啟動MZT,就會引發(fā)細胞周期停頓,導(dǎo)致發(fā)育延遲。
目前對m6A的轉(zhuǎn)錄組研究方法為MeRIP-seq(mRNA甲基化測序)的方法,其原理是通過m6A特異性抗體對細胞內(nèi)具有m6A修飾的RNA片段進行免疫共沉淀,將富集下來的RNA片段進行高通量測序。結(jié)合生物信息學(xué)分析,即可在轉(zhuǎn)錄組范圍內(nèi)對m6A修飾進行系統(tǒng)研究。
參考文獻:Zhao B S, Wang X, Beadell A V, et al. m(6)A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition.[J]. Nature, 2017, 542(7642):475.




















