互作轉(zhuǎn)錄組研究結(jié)核菌-宿主應(yīng)答差異
2017-12-13
病原菌需要迅速調(diào)整其毒性和適應(yīng)癥,以防止被宿主消滅。到目前為止,關(guān)于潛在適應(yīng)過程的驅(qū)動(dòng)致病機(jī)制大多是都是體外研究實(shí)驗(yàn),從而忽略了宿主誘導(dǎo)刺激對(duì)入侵病原體作用的真正復(fù)雜性。在本研究中,我們開發(fā)了一種實(shí)驗(yàn)方法,可以同時(shí)監(jiān)測(cè)宿主和侵染的細(xì)胞外病原體的轉(zhuǎn)錄變化。利用該工具可以發(fā)現(xiàn)許多與炎癥、穴位反應(yīng)、凝血活動(dòng)和過渡金屬離子隔離有關(guān)的宿主轉(zhuǎn)錄本的改變,并強(qiáng)調(diào)免疫反應(yīng)主要是浸潤中性粒細(xì)胞和混合TH17 / TH1反應(yīng)。因此,病原體的反應(yīng)主要是為了抵抗吞噬細(xì)胞的攻擊。肺結(jié)核菌上調(diào)了抗吞噬細(xì)胞III分泌系統(tǒng)(T3SS)的基因的表達(dá),并誘導(dǎo)了對(duì)抗中性白細(xì)胞誘導(dǎo)離子剝奪、自由基應(yīng)激和營養(yǎng)抑制的功能。對(duì)毒力影響最大的是碳儲(chǔ)存調(diào)節(jié)劑(Csr)系統(tǒng)的丟失,對(duì)宿主細(xì)胞T3SS表達(dá)上調(diào)至關(guān)重要。綜上所述,互作轉(zhuǎn)錄組測(cè)序?yàn)榘l(fā)現(xiàn)感染特異性刺激、誘導(dǎo)宿主和病原體反應(yīng)以及潛在的調(diào)節(jié)過程提供了有力的工具。
組織互作轉(zhuǎn)錄組結(jié)果
如圖1所示,(A)組織互作轉(zhuǎn)錄組結(jié)果顯示,雌性BALB/c小鼠口服感染2×108 cfu肺結(jié)核菌IP32953(感染)或1×PBS(未感染)。體外RNA-seq實(shí)驗(yàn)所用IP32953為25°C或37°C指數(shù)期或者平臺(tái)期的菌體。(B)基因組圈圖可視化顯示了IP32953染色體(NC_006155.1)和pYV質(zhì)粒(NC_006153.2)體外和體內(nèi)轉(zhuǎn)錄組數(shù)據(jù)。(C)從未受感染和感染的Peyer斑塊的RNA池中,對(duì)3重復(fù)的小鼠RNA-seq數(shù)據(jù)進(jìn)行了分析并制作熱圖。(D)主要成分(PC)分析均值為read數(shù)--來自結(jié)核菌IP32953核心基因組和pYV病毒質(zhì)粒體外和體內(nèi)轉(zhuǎn)錄組實(shí)驗(yàn)。
肺結(jié)核菌感染宿主反應(yīng)
如圖2所示, (A)DESeq2分析未受感染和受感染的Peyer的斑塊RNA獲得的火山圖。(B)前50富集和前50減少的宿主熱圖。(C)免疫介導(dǎo)的外部組織因子依賴性凝血級(jí)聯(lián)基因的活化。在IP32953的侵染過程中,受感染的Peyer斑塊中,外來的凝血級(jí)聯(lián)轉(zhuǎn)錄本已被高度濃縮。纖維蛋白凝塊通常是由細(xì)菌抗原(如LPS)誘導(dǎo)的,以誘捕入侵細(xì)菌并阻止進(jìn)一步的傳播。此外,通過中性細(xì)胞特異性膠原酶MMP8刺激外源性凝血通路,激活TFPI的組織因子。
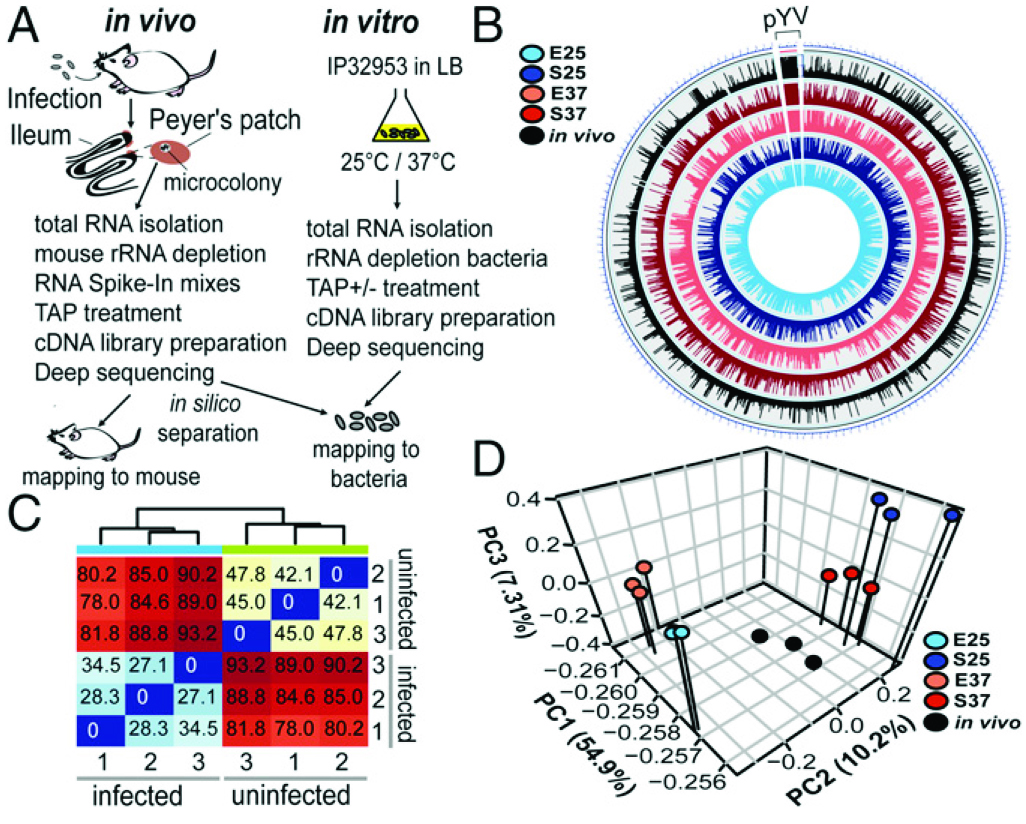
圖1組織互作轉(zhuǎn)錄組流程和報(bào)告。
毒力因子表達(dá)及宿主細(xì)菌應(yīng)激反應(yīng)
如圖3所示,細(xì)菌基因表達(dá)分析揭示了特定感染的代謝適應(yīng)性。(A)維恩圖展示了細(xì)菌侵染后的小鼠顯著改變的轉(zhuǎn)錄本(log2fc≥1;log2fc≤?1;P≤0.05);與4種體外實(shí)驗(yàn)結(jié)果相比,體內(nèi)實(shí)驗(yàn)有283個(gè)轉(zhuǎn)錄本發(fā)生了顯著變化,包括在一個(gè)條件下富集的和在另一種情況下衰竭。在這283個(gè)轉(zhuǎn)錄本中,116個(gè)被持續(xù)富集,62個(gè)在體內(nèi)持續(xù)衰竭。(B)在感染期間嚴(yán)格富集 (紅色)或衰竭(藍(lán)色)的選定細(xì)菌的基因熱圖。(C)結(jié)核桿菌中心碳代謝。在體外和體內(nèi)培養(yǎng)的細(xì)菌中,轉(zhuǎn)錄模式有了顯著變化。在感染過程中,富集的轉(zhuǎn)錄本用紅色表示,而減少的轉(zhuǎn)錄本用藍(lán)色表示。
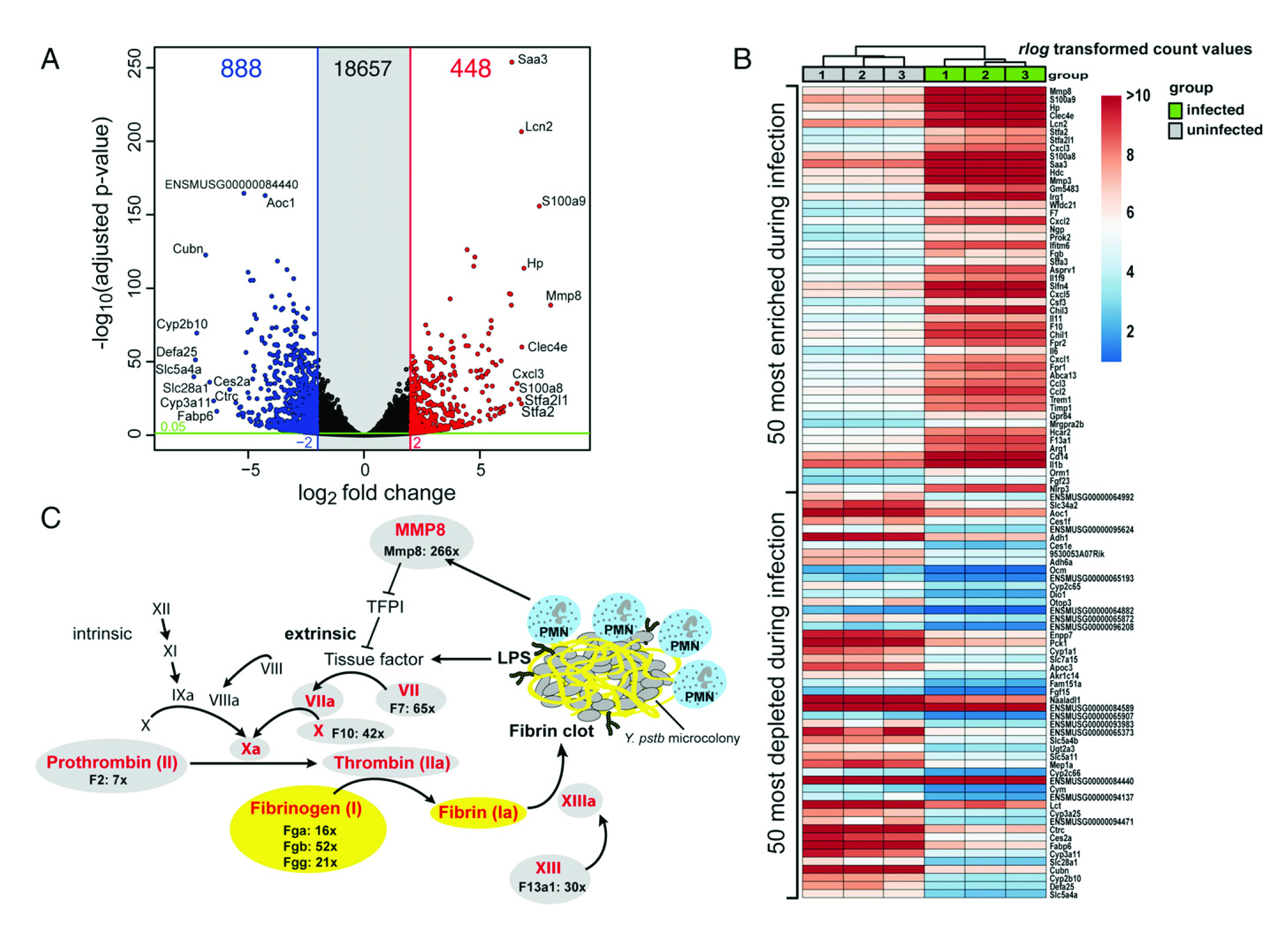
圖2宿主對(duì)肺結(jié)核菌的轉(zhuǎn)錄反應(yīng)
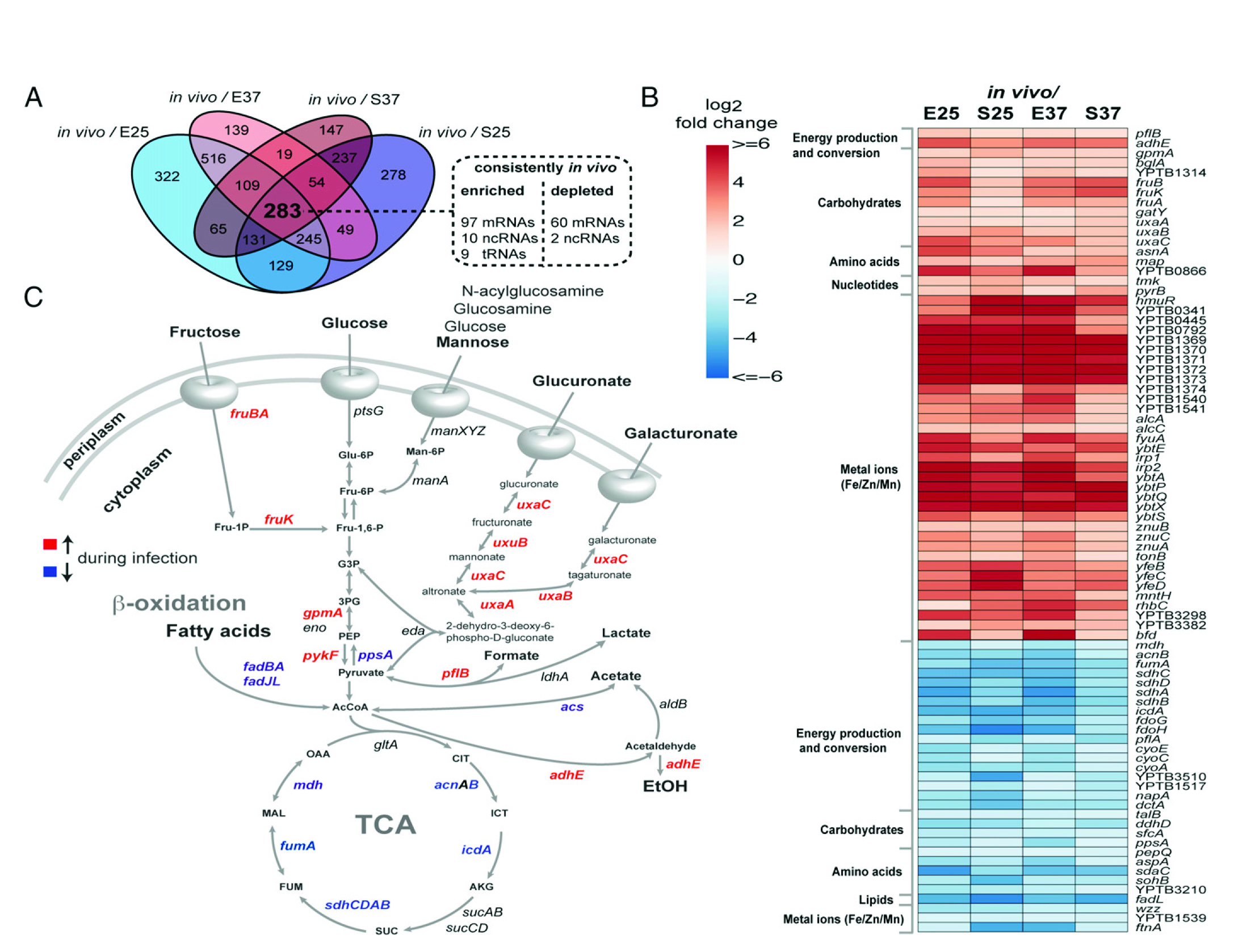
如圖3細(xì)菌基因表達(dá)分析
淋巴組織的生長改變了保守的代謝ncRNAs的數(shù)量
圖4顯示,高度保守的細(xì)菌ncRNAs有助于發(fā)病。(A)反式編碼的細(xì)菌ncRNAs的富集的熱圖(紅色)和衰竭的熱圖(藍(lán)色)。 (B) sgrS、ryhB1、ryhB2和glmY loci的rna - seq分析的read覆蓋率說明。數(shù)據(jù)按照惟一比對(duì)的reads進(jìn)行了規(guī)范化。(C)結(jié)核菌野生型IP32953和同基因的ΔsgrS在LB培養(yǎng)基培養(yǎng)37°C,添加和未添加αMG 1%,磷酸葡萄糖脅迫。(D)兩個(gè)獨(dú)立的BALB/c小鼠組(每組2×n = 5)口服等量混合感染107 cfu IP32953(WT)和同基因的突變YPIP56(ΔsgrS/ryhB1/ryhB2/glmY)。在Peyer斑塊(PPs)中,腸系膜淋巴結(jié)(MLNs)和3天后器官的組織樣本被添加不同的接頭。
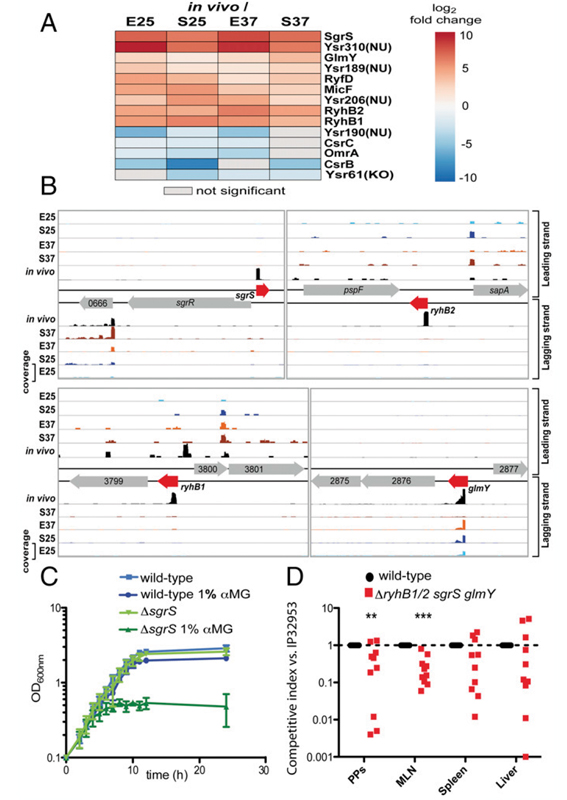
圖4 高度保守的細(xì)菌ncRNAs
圖5所示,CsrA對(duì)T3SS的毒力和介導(dǎo)其表達(dá)至關(guān)重要。(A)小鼠口服感染2×108 cfu結(jié)核菌野生型YPIII或同基因的ΔcsrA菌株。在兩個(gè)獨(dú)立的實(shí)驗(yàn)中每組每個(gè)基因型老鼠2×10只,實(shí)驗(yàn)12 d監(jiān)測(cè)老鼠的存活率。(B)BALB/c小鼠口服感染107 cfu結(jié)核菌YPIII WT或同基因的ΔcsrA菌株。小鼠感染3 d后被殺,確定均質(zhì)Peyer斑塊(PPs)、腸系膜淋巴結(jié)(MLNs)、肝臟和脾臟內(nèi)的細(xì)菌數(shù)目。數(shù)據(jù)來自兩個(gè)獨(dú)立實(shí)驗(yàn)(2×5老鼠/組)。(C)TCA-沉降上層清液或(D)Westernblots:全細(xì)胞提取物的結(jié)核菌YPIII WT和同基因的ΔcsrA菌株和沒有補(bǔ)充csrA +質(zhì)粒pAKH56生長在37°C缺少Ca2 +的培養(yǎng)基。(C)考馬斯亮藍(lán)染色或(D)Western blotting鑒定。核酸相關(guān)蛋白H-NS被用作陰性對(duì)照。
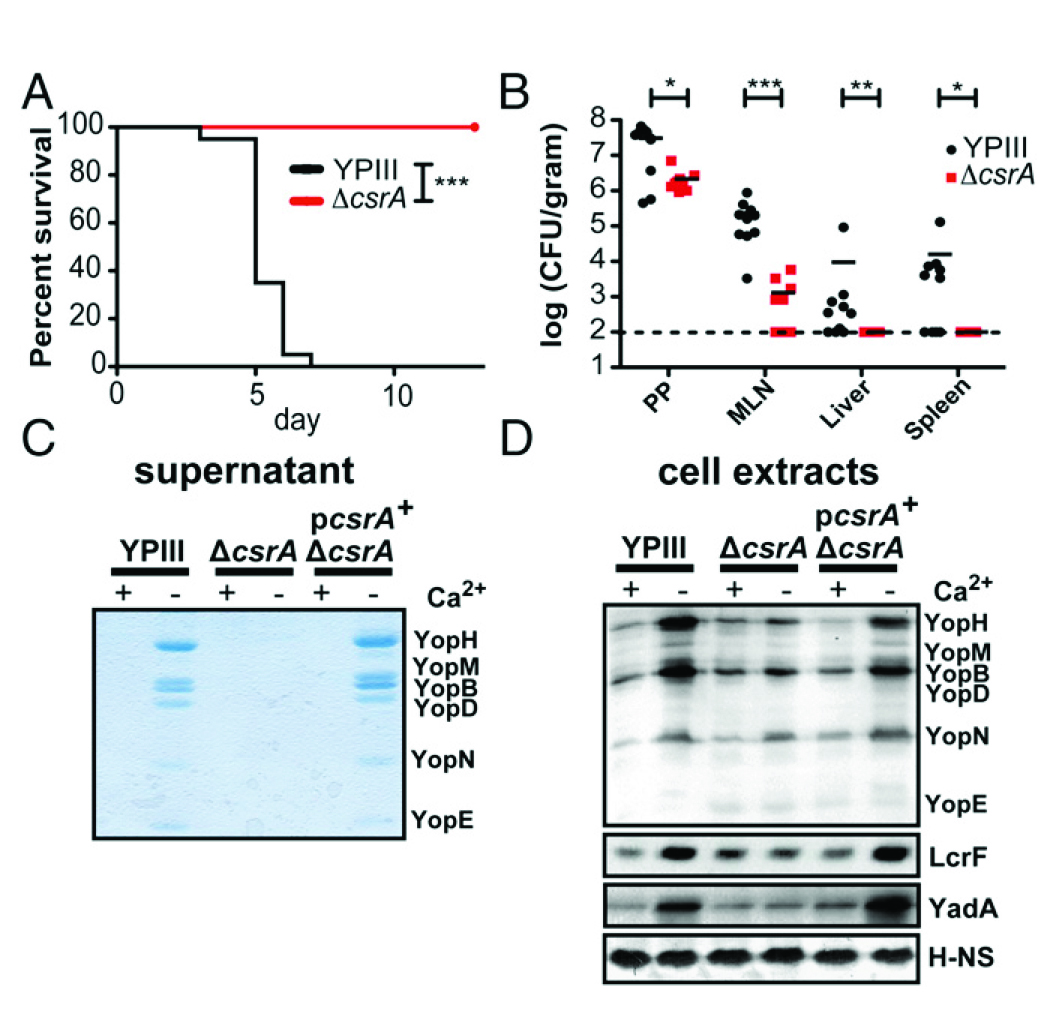
圖5 表型實(shí)驗(yàn)
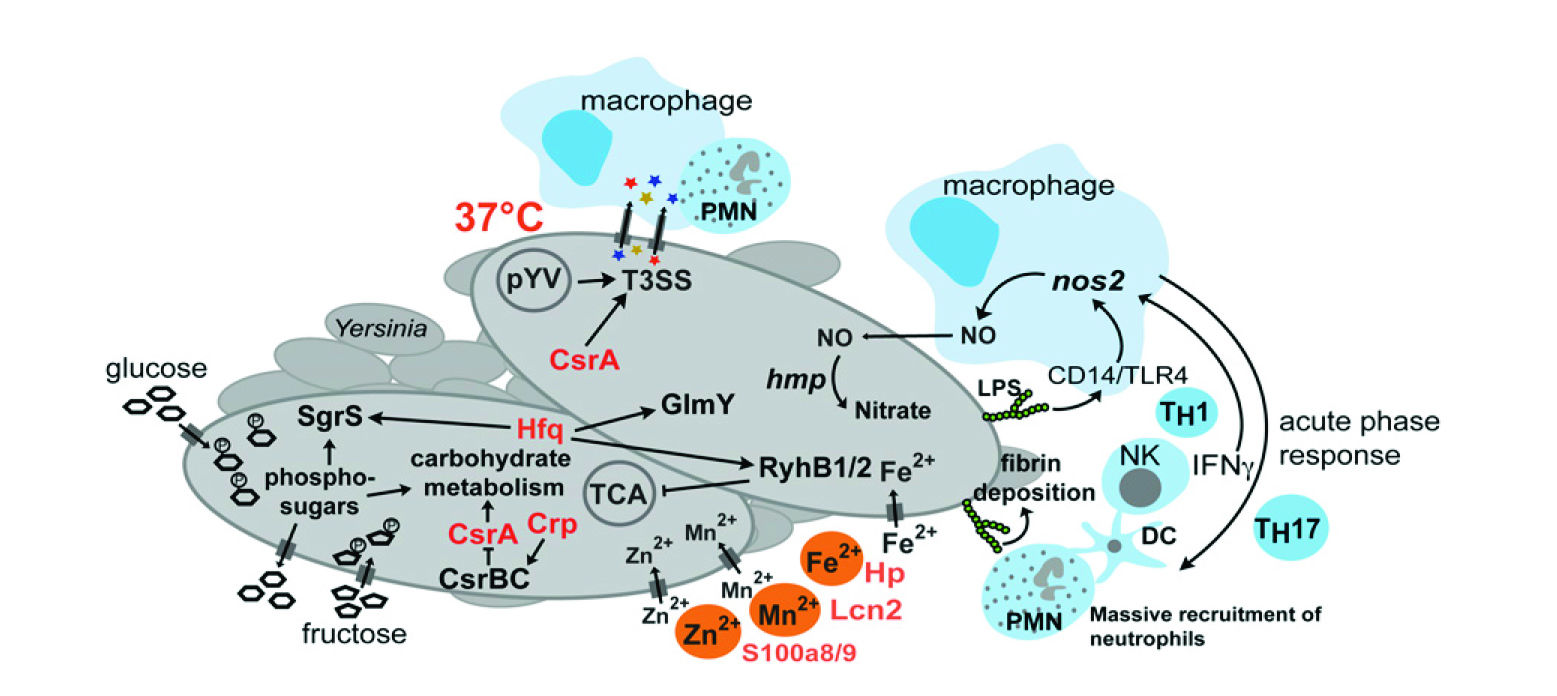
圖6 結(jié)核菌-宿主轉(zhuǎn)錄組變化
如圖6所示,結(jié)核菌-宿主轉(zhuǎn)錄組的特定感染變化。在Peyer斑塊的侵染過程中,對(duì)結(jié)核菌和宿主的復(fù)雜轉(zhuǎn)錄本適應(yīng)性的示意圖。最受影響的炎癥和急性期反應(yīng),代謝和毒力的適應(yīng),并識(shí)別不同的細(xì)菌核素從而影響發(fā)病機(jī)制。
綜上所述,以上結(jié)果證明了組織互作轉(zhuǎn)錄組方法的有用性,可以得出與病原體生物學(xué)和宿主反應(yīng)相關(guān)的有意義的結(jié)論。通過免疫細(xì)胞遷移和細(xì)菌致病機(jī)理的數(shù)據(jù)整合獲得的RNA剖面信息可以深入研究復(fù)雜的宿主-病原體相互作用網(wǎng)絡(luò)和發(fā)病機(jī)理。它進(jìn)一步創(chuàng)造了識(shí)別控制病原體侵染的關(guān)鍵過程的可能性,并增加了識(shí)別抗生素治療目標(biāo)的潛力。




















